![]()
CHAPTER ONE
Introduction
The fields of historical biogeography and ecological biogeography have long been paradoxically disparate and distant from one another, with different terminologies, different concepts, and almost nonoverlapping sets of researchers. Ecological biogeography focuses on spatial pattern in the composition and functioning of ecological communities, while historical biogeography attempts to reconstruct the history of areas and their biotas. Although some recent steps have narrowed gaps a bit, the two fields have long been quite distinct and disconnected, and spatial understanding of biodiversity has suffered as a consequence.
Differences between the two biogeographies are manifold: certainly, spatial scale is an important one, with most of ecological biogeography focusing at regional scales and most of historical biogeography at continental or even global scales. Another important difference is in treatment of temporal dimensions, with ecological biogeography focused chiefly over time spans that are geologically instantaneous (i.e., in the present), but historical biogeography looking from the present back over evolutionary time, sometimes many millions of years. Although not without significant exceptions (e.g., MacArthur 1972), these homonymous fields both have important insights to offer regarding the geography of biodiversity, yet have developed in large part independently until quite recently.
In recent years, however, an emerging body of work has begun to bridge between the two, building toward a more synthetic biogeography. Ecologists looking over broader spatial extents and into history, and systematists thinking about environmental dimensions and interactions among species, have come to understand that species' distributions are a function of phenomena from both realms. Detailed thinking regarding areas of distribution has also provided a fascinating reawakening of interest in another classic concept, that of the ecological niche. In effect, understanding areas of distribution of species in terms of their ecological requirements across multiple scales of space and time has provided an arena for a meeting of these two disparate disciplines. The impressive impact on both fields of just the first few years of this interaction suggests that their integration will have a bright future.
PRACTICALITIES
The past few years have witnessed substantial increases in availability of species' occurrence data, also termed “primary biodiversity data” or “presence data” or “occurrence data” (Soberón et al. 1996, Graham et al. 2004a, Soberón and Peterson 2004). This trend results from large-scale efforts to digitize and reference to geographic coordinates (“georeference”) the estimated 1-3 billion specimens held in world museums and herbaria (Chalmers 1996, Krishtalka and Humphrey 2000), as well as efforts to improve access to large observational data stores, at least for certain taxonomic groups (chapter 5). Presence data, as we will see in coming chapters, form the basis for most efforts to estimate ecological niches. Publicly accessible Internet portals now allow access to on the order of 300 million primary biodiversity data records (Edwards 2004).
Information regarding environmental variables is now similarly abundant. Petabytes (i.e., millions of gigabytes) of environmental information about climate, topography, soils, oceanographic variables, vegetation indices, land-surface reflectance, and so on, are available across almost the entire planet, and at increasingly finer resolutions (chapter 6). These datasets are being generated by agencies such as the European and U.S. space agencies, by the United Nations, by university researchers (e.g., Hijmans et al. 2005), and by many national institutions (e.g., CONABIO 2009, INPE 2009, NRSC 2009).
Finally, powerful software allowing estimation of both areas of distribution and theoretical objects related to niches has been implemented. The work of pioneers like Grinnell, Hutchinson, and Austin suddenly became thriving research areas linking ecological and historical dimensions of biogeography. In particular, these tools enable what has been termed “species distribution modeling” (SDM; Guisan and Zimmermann 2000, Hirzel et al. 2002, Guisan and Thuiller 2005, Araújo and Guisan 2006), as well as the related (but by no means equivalent) endeavor called “ecological niche modeling” (ENM; Peterson et al. 2002d, Soberón and Peterson 2005, Soberón 2007). These fields—the subject of this book—center on application of niche theory to questions about real and possible spatial distributions of species in the past, present, and future. In a very real sense, the availability of large quantities of data, technological developments like geographic information systems (GIS), and several computational tools are enabling a multitude of applications that are not only of biological importance, but that also can often be of extreme practical utility.
Nevertheless, many carefully pondered decisions are necessary before it is possible to turn these data and tools into interesting analyses and useful knowledge with full scientific rigor. Many crucial methodological issues remain to be explored and resolved, like the types of variables to be included, whether to use them in raw or transformed forms, whether to reduce dimensionality prior to analysis, what spatial and temporal resolution to use, and how best to assess model performance; all of these points are questions affecting niche modeling exercises. The answers to these practical questions depend on a rigorous conceptual framework. After a period of development in which conceptual and methodological rigor took a back seat to rapid development of software and data resources, the time has come to take stock of the advances and propose a conceptual reorganization. All of this thinking is the subject of this book.
THIS VOLUME
We offer this volume as a first synthesis of concepts in this emerging field. In spite of hundreds of research contributions and increasing numbers of reviews and commentaries, no rigorous and quantitative conceptual framework and synthesis has been presented. This lack of synthesis is nowhere more notable than in the debate between groups of researchers using the same tools to address the same questions, yet employing—whether knowingly or not—very distinct conceptual frameworks in the development of their analyses and conclusions. Such misunderstandings can be avoided, if a common language and thinking framework are available. This book represents the crystallization of years of thinking and work by a diverse suite of coauthors, all interested in the ecological, geographic, and evolutionary dimensions of geographic distributions of species.
We do not intend this book to serve as an exhaustive review of the burgeoning literature on ecological niches and geographic distributions. Quite simply, just in the time in which we have been preparing this manuscript, hundreds of new papers have been published, making the idea of an exhaustive review a moving target that is probably impossible to achieve. Moreover, recent publication of a book by J. Franklin (2010), Mapping Species Distributions, does a commendable job of reviewing and synthesizing the vast literature on this topic. Our approach is more conceptual: we aim to offer a body of terminology and schemes by which to understand and discuss phenomena of distributional ecology; a common language is badly needed in a field so rife with ill-defined jargon and loosely defined terms.
This book focuses on the complex relationships between ecological niches and geographic distributions of species, both across space and (perhaps to a lesser degree) through time. We provide a conceptual overview, which we hope will be of broad interest to researchers interested in diverse aspects of ecology, biogeography, and other related fields. However, we focus much of this book on how that conceptual framework links to the emerging fields of ecological niche modeling and species distribution modeling, both correlative approaches to understanding ecological niches and geographic distributions.
We do not make any broad attempt to provide a similar overview or deep understanding of process-based, physiological approaches to estimating ecological niches that other researchers are exploring—rather, we see the two approaches as complementary. The process-based approaches show considerable promise, and in some situations offer the only view possible into the fundamental ecological niches of species. However, such approaches remain in early stages of development and exploration. This book focuses on the correlative approaches, which we see as most broadly applicable to diverse questions regarding the ecology and geography of biodiversity phenomena.
The reader should not imagine that this process of finding a common language has been easy. Indeed, even among the seven authors of this book, who have worked together for years, some strong differences of opinion still exist regarding terminologies and concepts. We have, however, achieved a high degree of concordance, and have been willing to look past our colleagues' different views in striving for synthesis. Many of the insights that emerged from these debates concerned the relevance of ideas central to each of our respective backgrounds and skill sets, including integration of ideas from field biology, morphology, systematics, genetics, theoretical ecology, evolutionary biology, statistics, climatology, and geospatial science.
The result is this volume. We begin with a conceptual framework for thinking about and discussing the distributional ecology of species, which has involved considerable exploration of the field of population ecology, and has required revisiting several “sacred” texts, such as the fundamental early works of Hutchinson and Grinnell. A second section addresses the data and tools that have been marshalled in the early development of this field. We avoid carefully the temptation to review and assess specific software tools, as we consider these to be transitory and less important than the base concepts. It is much more important that the field have a consistent terminology and thinking framework than to “know” that such and such program is the “best.” (Besides, as the reader will see in the chapters that follow, what appears to be the “best” frequently is not what it appears, and is certainly context- and scale-dependent.) In the final section, we provide a relatively brief overview of real-world situations to which these tools have been applied, to illustrate the promise of this new field. Our hope is to move the discourse in this field to a new level, once a common platform of ideas is established.
![]()
PART I
THEORY
![]()
CHAPTER TWO
Concepts of Niches
It has often been pointed out that the term “niche” disguises several concepts under a single label (Whittaker et al. 1973, Colwell 1992, Leibold 1995, Chase and Leibold 2003, Odling-Smee et al. 2003). Some authors, perhaps overwhelmed by the broad variety and subtle shades of meaning assigned to the word, have advised that “niche is perhaps a term best left undefined” (Bell 1982). We disagree: in science, arguments benefit from precise and consistent usage of key concepts; otherwise, clear thinking is hindered. Using the same word to refer to different ideas leads to confusion, as the picturesque history of the word “niche” amply demonstrates. Besides, different senses of niche are appropriate to deal with different biological problems. As a consequence, several niche concepts exist, and our first task is thus to clarify and specify which concept we will be using and why. We will choose terminology and ideas best suited to the problem in which we are interested—namely, that of estimating and understanding areas of distribution of species.
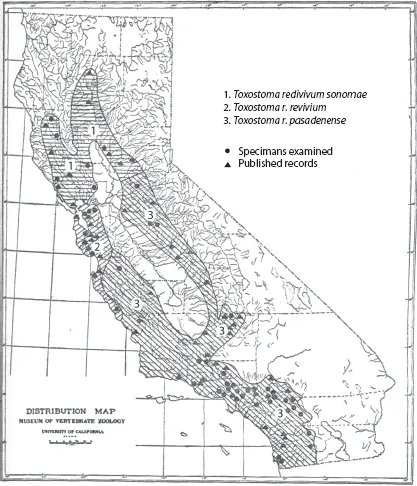
One reason why “niche” has acquired a veritable bush of meanings is that, since the first time it was used, ecologists have applied the term to analyze a very complex question: what combinations of environmental factors allow a species to exist in a given geographic region or in a given biotic community, and what effects does the species have on those environmental factors? Not only does the preceding statement refer to an intrinsically complex set of problems, but several of its terms can be interpreted and measured in a variety of ways. Moreover, the concept has been used at both geographic and local scales, most often assuming that the ensuing complications and differences should be obvious. For example, Grinnell (1917), studying the niche of the California Thrasher (Toxostoma redivivum) in relation to its area of distribution (this idea will be discussed later as the “existence” of the species), meant by the term niche the thrasher's climatic and habitat requirements (the environmental factors) expressed geographically (figure 2.1). In a contrasting interpretation, Elton (1927) viewed the niche as the functional role of an animal on a community (its local effects): the existence was taken for granted, but the emphasis was placed on the “impacts.” These two early views of niche illustrate one of the main causes of ambiguity of concept: stress on requirements at geographic scales versus stress on impacts at local scales (Chase and Leibold 2003).
FIGURE 2.1. Distribution of the California Thrasher (Toxostoma redivivum) in California, from Grinnell (1917). Approximate distributional limits of the three subspecies are shown with different shadings, and occurrences are shown as dots (for specimens) or triangles (for published records).
After reviewing the history of niche concepts, Chase and Leibold (2003) concluded that much of the confusion surrounding the term results because “previous authors have not consistently distinguished between the responses of organisms to their environment and the effects of organisms on their environment.” Indeed, this difference has both deep implications about the actual mathematical form of a multivariate niche definition and serious operational consequences, since certain variables related to requirements can be measured easily, whereas most variables related to impact require ad hoc experimental efforts.
Although attempting an exhaustive classification of niche concepts based on the preceding ideas would be interesting, that task is not our purpose in this chapter. Rather, we intend to propose a formal and operational definition of a particular niche concept (which is naturally related to the problem of estimating areas of distribution), offer approaches to characterize and measure it, and use it as a conceptual and terminological basis for describing and understanding much of the related practices of ecological niche modeling and species distribution modeling (Peterson 2006c). In this chapter, we explain the reasons for our choice of emphases, leading to a particular meaning and usage of niche. To accomplish this goal, we review briefly the themes most important in understanding niche concepts, highlighting the meanings most appropriate to the purpose of this book.
MAJOR THEMES IN NICHE CONCEPTS
Recall Hutchinson's (1957) definition of the fundamental niche of a species: a hypervolume of environmental variables, “every point of which corresponds to a state of the environment which would permit the species to exist indefinitely.” Most differences in niche concepts depend on the formulation and relative importance given to three interrelated points, considered in turn later: (1) the meaning of “exist indefinitely,” (2) what kinds of variables constitute the hypervolume, and (3) the nature of feedback loops between a species and the variables composing the hypervolume. Our definitions of niche are based on operational specifications of the preceding three points.
THE “EXISTENCE” OF A SPECIES
As highlighted by Holt (2009), several ambiguities complicate the term “existence” of the species, as used in niche definitions. From a population ecology point of view, a species may exist for a period of time t0 < t < tl in a place g of the world, if its total instantaneous growth rate dxg(t)/dt is on average nonnegative during t0 < t < tl (Vandermeer 1972, Maguire 1973, Hutchinson 1978). However, and disregarding stochastic factors and evolutionary change, the instantaneous growth rate is composed (see next chapter) of (1) an intrinsic growth rate rg that is by definition density-independent, (2) density-dependent factors and the results of interactions with other species, and (3) the population structure that determines arrival of dispersing individuals to the locality g and that may allow existence of the species in places where rg < 0 (sink populations; Pulliam 1988 and 2000).
It is clear that quite-different combinations of the preceding factors may lead to a species “existing” [i.e., dx(t)/dt > 0] or not in a given region over a given period of time. Holt (2009) pointed out that some of the major issues requiring clarification are (1) the existence of Allee effects (reduced per-capita growth rate at small population densities) that will make some regions where rg > 0 impossible to invade because the flow of migrants is below the Allee threshold; (2) the possibility that, in regions of high instantaneous growth rate, population growth will lead to irreversible alterations of the envir...