Miriam Zelditch, Donald Swiderski, H. David Sheets
This is a test
This is a test
Compartir libro
488 páginas
English
ePUB (apto para móviles)
Disponible en iOS y Android
eBook - ePub
Geometric Morphometrics for Biologists
A Primer
Miriam Zelditch, Donald Swiderski, H. David Sheets
Detalles del libro
Vista previa del libro
Índice
Citas
Información del libro
The first edition of Geometric Morphometrics for Biologists has been the primary resource for teaching modern geometric methods of shape analysis to biologists who have a stronger background in biology than in multivariate statistics and matrix algebra. These geometric methods are appealing to biologists who approach the study of shape from a variety of perspectives, from clinical to evolutionary, because they incorporate the geometry of organisms throughout the data analysis. The second edition of this book retains the emphasis on accessible explanations, and the copious illustrations and examples of the first, updating the treatment of both theory and practice. The second edition represents the current state-of-the-art and adds new examples and summarizes recent literature, as well as provides an overview of new software and step-by-step guidance through details of carrying out the analyses.
Contains updated coverage of methods, especially for sampling complex curves and 3D forms and a new chapter on applications of geometric morphometrics to forensics
Offers a reorganization of chapters to streamline learning basic concepts
Presents detailed instructions for conducting analyses with freely available, easy to use software
Provides numerous illustrations, including graphical presentations of important theoretical concepts and demonstrations of alternative approaches to presenting results
Preguntas frecuentes
¿Cómo cancelo mi suscripción?
Simplemente, dirígete a la sección ajustes de la cuenta y haz clic en «Cancelar suscripción». Así de sencillo. Después de cancelar tu suscripción, esta permanecerá activa el tiempo restante que hayas pagado. Obtén más información aquí.
¿Cómo descargo los libros?
Por el momento, todos nuestros libros ePub adaptables a dispositivos móviles se pueden descargar a través de la aplicación. La mayor parte de nuestros PDF también se puede descargar y ya estamos trabajando para que el resto también sea descargable. Obtén más información aquí.
¿En qué se diferencian los planes de precios?
Ambos planes te permiten acceder por completo a la biblioteca y a todas las funciones de Perlego. Las únicas diferencias son el precio y el período de suscripción: con el plan anual ahorrarás en torno a un 30 % en comparación con 12 meses de un plan mensual.
¿Qué es Perlego?
Somos un servicio de suscripción de libros de texto en línea que te permite acceder a toda una biblioteca en línea por menos de lo que cuesta un libro al mes. Con más de un millón de libros sobre más de 1000 categorías, ¡tenemos todo lo que necesitas! Obtén más información aquí.
¿Perlego ofrece la función de texto a voz?
Busca el símbolo de lectura en voz alta en tu próximo libro para ver si puedes escucharlo. La herramienta de lectura en voz alta lee el texto en voz alta por ti, resaltando el texto a medida que se lee. Puedes pausarla, acelerarla y ralentizarla. Obtén más información aquí.
¿Es Geometric Morphometrics for Biologists un PDF/ePUB en línea?
Sí, puedes acceder a Geometric Morphometrics for Biologists de Miriam Zelditch, Donald Swiderski, H. David Sheets en formato PDF o ePUB, así como a otros libros populares de Mathematics y Applied Mathematics. Tenemos más de un millón de libros disponibles en nuestro catálogo para que explores.
Owing largely to developments in measurement theory over the past two decades, there has been remarkable progress in morphometrics. That progress resulted from first precisely defining “shape” and then pursuing the mathematical implications of that definition. We therefore now have a theory of measurement. We offer a critical overview of the recent history of measurement theory, presenting it first in terms of exemplary data sets and then in more general terms, emphasizing the core of the theory underlying geometric morphometrics – the definition of shape. We conclude the conceptual part of this Introduction with a brief discussion of methods of data analysis. The rest of the Introduction is concerned with the organization of this book and where you can find more information about available software and other resources for carrying out the analyses.
Keywords
geometric morphometrics, shape analysis, landmark coordinates, shape and size
Shape analysis plays an important role in many kinds of biological studies. A variety of biological processes produce differences in shape between individuals or their parts, such as disease or injury, mutation, ontogenetic development, adaptation to local geographic factors, or long-term evolutionary diversification. Differences in shape may signal differences in processes of growth and morphogenesis, different functional roles played by the same parts, different responses to the same selective pressures, or differences in the selective pressures themselves. Shape analysis is an approach to understanding those diverse causes of morphological variation and transformation.
Sometimes, differences in shape are adequately summarized by comparing the observed shapes to more familiar objects such as circles, kidneys or letters of the alphabet (or even, in the case of the Lower Peninsula of Michigan, a mitten). Organisms, or their parts, are then characterized as being more or less circular, reniform, C-shaped or mitten-like. Such comparisons can be extremely valuable because they help us to visualize unfamiliar organisms or to focus attention on biologically meaningful components of shape. However, they can also be vague, inaccurate or even misleading, especially when the shapes are complex and do not closely resemble familiar icons. Even under the best of circumstances, we still cannot say precisely how much more circular, reniform, or C-shaped or mitten-like one shape is than another. When we need that precision, we turn to measurement.
Morphometrics is a quantitative way of addressing the shape comparisons that have always interested biologists. This may not seem to be the case because the morphological approaches once typical of the quantitative literature appeared very different from the qualitative descriptions of morphology; whereas the qualitative studies produce pictures or detailed descriptions (in which analogies figure prominently), morphometric studies usually produced tables with disembodied lists of numbers. Those numbers seemed so highly abstract that we could not readily visualize them as descriptors of shape differences, and the language of morphometrics also seemed highly abstract and mathematical. As a result, morphometrics seemed closer to statistics or algebra than to morphology. In one sense that perception is entirely accurate: morphometrics is a branch of mathematical shape analysis. The way that we extract information from morphometric data involves mathematical operations rather than concepts rooted in biological intuition or classical morphology. Indeed, the pioneering work in modern geometric morphometrics (the focus of this book) had nothing at all to do with organismal morphology; the goal was to answer a question about the alignment of megalithic “standing stones” like Stonehenge (Kendall and Kendall, 1980). Nevertheless, morphometrics can be as much a branch of morphology as it is a branch of statistics. It is that when the tools of shape analysis are turned to organismal shapes, illustrating and even explaining shape differences that have been mathematically analyzed.
The tools of geometric shape analysis have a tremendous advantage when it comes to these purposes: not only because it offers precise and accurate description, but also because it enables rigorous statistical analyses and serves the important purposes of visualization, interpretation and communication of results. Geometric morphometrics allows us to visualize differences among complex shapes with nearly the same facility as we can visualize differences among circles, kidneys and letters of the alphabet (and mittens).
In emphasizing the biological component of morphometrics, we do not discount the importance of its mathematical component. Mathematics provides the models used to analyze data, both the general linear models exploited in statistical analyses and the algebraic models underlying exploratory methods such as principal components analysis. Additionally, mathematics provides a theory of measurement that we use to obtain the data in the first place. It may not be obvious that any theory governs measurement because very little theory (if any) underlays traditional measurement approaches. Asked the question “What are you measuring?”, we could give many answers based on our biological motivation for measurement – such as (1) “functionally important characters”; (2) “systematically important characters”; (3) “developmentally important characters”; or (4) “size and shape”. However, when asked “what do you mean by “character” and how is that related either mathematically or conceptually to what you are measuring?” or if asked “what do you mean by “size and shape”?”, it was difficult to provide coherent answers. A great deal of experience and tacit knowledge went into devising measurement schemes, but that knowledge and experience had very little to do with any general theory of measurement. Rather than being grounded in a general theory of measurement, each study appeared to devise its approach to measurement according to the biological questions at hand, as guided by the particular tradition within which that question arose. There was no general theory of shape nor were there any analytic methods adapted to the characteristics of shape data.
Owing largely to developments in measurement theory over the past two decades, there has been remarkable progress in morphometrics. That progress resulted from first precisely defining “shape” and then pursuing the mathematical implications of that definition. We therefore now have a theory of measurement. Below we offer a critical overview of the recent history of measurement theory, presenting it first in terms of exemplary data sets and then in more general terms, emphasizing the core of the theory underlying geometric morphometrics – the definition of shape. We conclude the conceptual part of this Introduction with a brief discussion of methods of data analysis. The rest of the Introduction is concerned with the organization of this book and where you can find more information about available software and other resources for carrying out morphometric analyses.
A Critical Overview of Measurement Theory
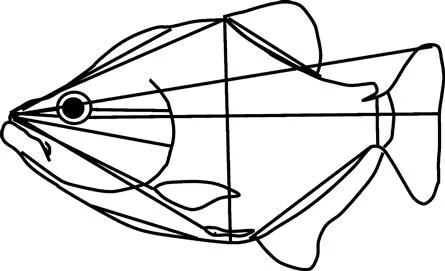
Traditionally, morphometric data were measurements of length, depth and width, such as those shown in Figure 1.1, based on a scheme presented in a classic ichthyology text (Lagler et al., 1962). Such a data set contains relatively little information about shape and some of it is fairly ambiguous. These kinds of data sets contain less information than they appear to hold because many of the measurements overlap or run in similar directions. What may be most obvious is that several measurements radiate from a single point so that their values cannot be completely independent; any error in locating that point affects all of these measurements. Such a data set contains less information than could have been collected with no greater effort because some directions are measured redundantly and many measurements overlap. For example, there are many measurements of length along the anteroposterior body axis and most of them cross some part of the head, whereas there are only two measurements along the dorsoventral axis and both are of post-cranial dimensions. In addition, because most of the measurements are long, it is difficult to localize shape differences to any region, such as any change in the proportions of the pre- and postorbital head or the position of the dorsal fin relative to the back of the head. Also, some of the information that is missing from this type of measurement scheme, but which is necessary for morphological analysis, concerns the spatial relationships among measurements. That information might be in the descriptions of the measurements, i.e. the line segments, but it is not captured by the data. The data consist solely of a list of observed values of those lengths. Finally, the measurements may not sample homologous features of the organism, making it difficult to interpret the results. For example, body depth can be measured by a line extending between two well-defined points (e.g. the anterior base of the dorsal fin to the anterior base of the anal fin), but it can also be measured wherever the body is deepest, yielding a measurement of “greatest body depth” wherever that occurs. That second measurement of depth might not be comparable anatomically from species to species, or even from specimen to specimen, so it provides almost no useful information (except for maximal depth). Considering these many limitations of traditional measurements, it is clear that the number of measurements greatly overestimates the amount of shape information that is actually collected.
Figure 1.1 Traditional morphometric measurements of external body form of a teleost, adapted from the scheme in Lagler et al., 1962.
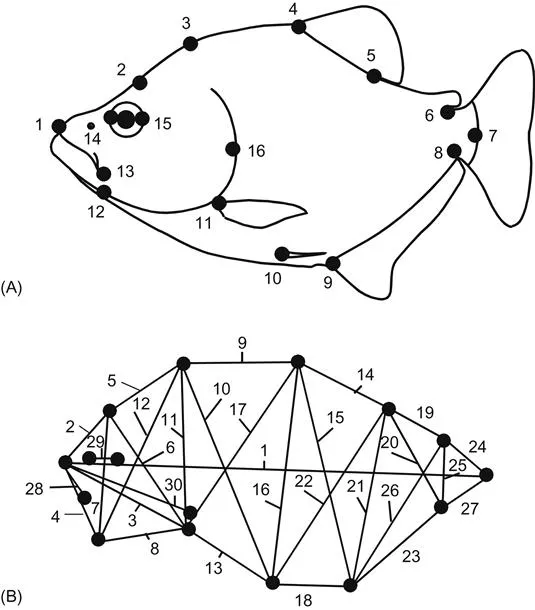
The classical measurement scheme can be greatly improved without altering its basic mathematical framework, by the box truss (Figure 1.2) – a scheme developed by Strauss, Bookstein and colleagues (Strauss and Bookstein, 1982; Bookstein et al., 1985). This set of measurements samples more directions of the organism, the measurements are more evenly spaced, and there are also many short measurements. Moreover, all the endpoints of the measurements are biologically homologous anatomical loci – landmarks. But even though the truss is a clear improvement over classical measurement schemes when it comes to describing shape differences, the result is still just a list of numbers (i.e. the lengths of the truss elements), with all the attendant problems of visualization and communication.
Figure 1.2 Truss measurement scheme of external body form of a teleost: (A) well-defined endpoints of measurements; (B) a selection of 30 lengths, arranged in a truss.
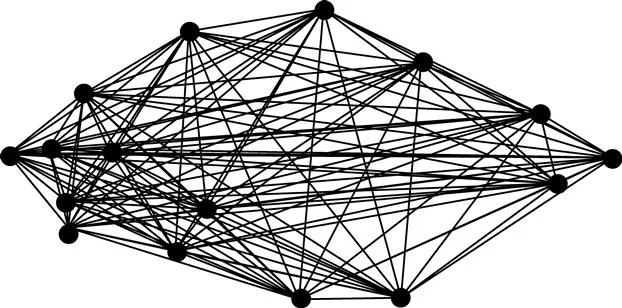
One general problem shared by both those measurement schemes is that they fail to collect all the information available from the endpoints of the measurements. The truss scheme shown in Figure 1.2 contains 30 measurements, but 30 is only a fraction of the 120 that could be made among the same 16 landmarks (Figure 1.3). Of course, many of the 120 are redundant, and several of them span large regions of the organism, making it difficult to localize where changes occur. Additionally, we would need extraordinarily large samples in order to test hypotheses about shape and the results would be incredibly difficult to interpret because there would be 120 pieces of information (e.g. regression coefficients, principal component loadings) for each trend or difference. Analyzing all 120 requires specialized methods beyond the scope of this book (see Lele and Richtsmeier, 1991, 2001; Richtsmeier and Lele, 1993). We might be tempted to cull the 120 measurements, retaining only those that seem most likely to be informative but, until we have done the analysis, we cannot know which can be safely culled. Clearly, we need another way to get the same shape information as the 120 measurements, but without the excessive redundancy.
Figure 1.3 All 120 measurements between endpoints defined by the 16 landm...