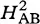This book is unique; the factual content and ideas it expounds are only just beginning to be touched upon in standard texts. Protein Electron Transfer is a major collaborative effort by leading experts and explores the molecular basis of the rapidly expan
The chemiosmotic hypothesis describes the central feature of biological energy transduction as the creation of a transmembrane proton gradient from redox potential energy of photosynthesis and respiration (Mitchell, 1961). The gradient is built up by a series of guided electron transfers within and between membranous proteins that selectively move electrons and protons without compromising the insulation of the membrane. Energy transduction relies on the integration of three mechanisms: (i) diffusional motion of protons and electron carriers between specific sites on the major redox complexes; (ii) coupling of electron and proton motion at sites designed to control the bond making and breaking chemistry; and (iii) long-range electron transfer through the insulating medium between sites.
This last mechanism relies on electron tunnelling, a phenomenon that was clearly shown to be important in biology by early experiments of DeVault and Chance (1966). Light-activated electron transfer in the photosynthetic bacterium Chromatium vinosum was shown to have a relatively constant millisecond rate over a wide cryogenic temperature range down to liquid helium temperatures, the hallmark of a long distance electron tunnelling reaction. Since this time, biological electron transfer has benefited from advances in the physical theory of tunnelling and, in return, biology has provided a fine experimental field for testing theory (DeVault, 1980).
Biological systems appear astonishingly complex. Even small protein units have several thousands of atoms integrated into convoluted structures. Attempts to build a complete theoretical description of the behaviour of these systems are forced immediately into drastic simplifications (Friesner, 1994; Newton, 1991). For the small percentage of electron transfer complexes where we fortunately have crystals in which X-ray diffraction provides some approximation of one possible geometry, no amount of available computing ability can support ab initio calculations of all the wave functions involved. Instead various approximations and functional are needed to begin a simulation. In this sort of environment, simple models and formalisms have an important role, especially if such pictures integrate a large number of observations and provide a much needed set of guidelines with which to test electron transfer theory, stimulate predictive insight into biological function and plan further experiments.
1.2 Fundamental tunnelling theory
1.2.1 Simple electron tunnelling through a barrier
The simplest picture of electron tunnelling involves the localization of an electron on a donor in a narrow potential well. Surrounding the donor is an electrically insulating region which represents a barrier to the electron. Classically the electron does not have enough energy to surmount the barrier yet, in the quantum mechanical view, it is possible to find the electron with some small but finite probability inside the barrier region. That is, the wave function describing the electron penetrates the insulating barrier. Indeed, the wave function penetrates far enough to reach the potential well of an acceptor, and thus there is a finite probability that an electron localized on the donor will be found at the acceptor. In the simple case of narrow donor and acceptor wells of equal depth, and a uniform intervening barrier of uniform height V above the wells, the penetration of the electron will fall off exponentially with distance (R) and will be given by Equation 1.1 (Gamow, 1928):
where
represents the square of the electronic coupling of the donor and acceptor states which is related to the square of the integrated overlap of the donor and acceptor electronic wave functions, m is the mass of the electron and ħ is Planck’s constant/2π. In a fortunate coincidence of dimensional units, the exponential coefficient (β) in units of Å−1 will be approximately the square root of the barrier height in eV.
There is no temperature term in this expression. This is a ‘deep-well’ picture, in which the barriers sensed by the electron are measured in electron volts, much larger than physiological thermal energies of about 25 meV. This tunnelling phenomenon contrasts with a classical transition state view in which thermal energy is required to cross over, as opposed to pass through, a barrier, thereby revealing an Arrhenius activation energy. This absence of a significant activation energy barrier at cryogenic temperatures, together with the relatively slow millisecond reaction rate, suggested long distance electron tunnelling to DeVault and Chance (1966).
1.2.2 Fermi’s golden rule
To relate the tunnelling theory to observation, we must be able to derive an electron transfer rate. However, in the simple quantum mechanical view just described, an electron will penetrate the barrier to the acceptor well but then return through the barrier back to the donor in a resonant process that depends on the distance between and the relative energy of the wells. In any real biological system there will be a range of acceptor wells of slightly different depths (i.e. there is a density of acceptor states) which break up the resonance and lead to a time dependence which can define an electron transfer rate. The corresponding mathematical description of this electron transfer rate (ket; Equation 1.2) is the fundamental equation of non-adiabatic electron transfer theory, Fermi’s golden rule:
FC refers to the Franck–Condon weighted density of states. As in spectroscopy, the Franck–Condon term refers to an electron jumping from one quantum state to another in a time too short for the nuclei to respond; a large FC term corresponds to a favourable geometric overlap of the nuclear wave functions of the reactant and product. Thus Equation 1.2 also serves to separate the dependence of the electron transfer rate into electronic and nuclear terms. This equation is applicable only to cases in which the donor and acceptor are relatively well separated and distinct, in other words long distance electron transfer. It is critical to remember that, in cases where the donor and acceptor are close enough to interact strongly, electron transfer is adiabatic and this simple description has to be abandoned; prediction of electron transfer rates becomes much more difficult.
1.2.3 Marcus theory and nuclear geometries
A particularly straightforward interpretation of the Frank–Condon term has been provided by Marcus (Marcus, 1956; Marcus and Sutin, 1985). In this view (Figure 1.1), the nuclei of the reactant and its immediate environment are represented by a single simple harmonic oscillator potential along a reaction coordinate. The minimum of this potential represents the equilibrium geometry of the reactant. The product is represented by a similar harmonic potential with a minimum that is displaced along the reaction coordinate to the equilibrium geometry of the product, and lowered in potential by AG, the free energy of the electron transfer reaction. The change in geometry upon electron transfer is characterized by the reorganization energy (A), defined as the energy that must be added to a reactant and its environment to distort its equilibrium geometry into the equilibrium geometry of the product without, however, allowing the electron to be transferred. It includes, for example, the energy required to rearrange the solvent dipoles from a geometry that electrostatically favours the electron on the donor, to a geometry that favours the electron on the acceptor.
In this non-adiabatic view, the reactant can be viewed as moving back and forth along the parabola surface, passing through the region of intersection with the product parabola many times before the weak coupling permits electron transfer to take place. In contrast, in an adiabatic reaction, the reactant and product are so strongly coupled that motion in this region can carry the reactant directly to the product. In the adiabatic limit, the bottoms of the two roughly parabolic wells are joined into a single, more complex surface which is separated, or split, from the remnants of the upper parabolic surfaces by the large electronic coupling, 2HAB.
Figure 1.1. In a Marcus description, all the nuclear motion of the reactant and the surrounding environment are approximated as a single, simple harmonic oscillator potential (upper parabola on left). The equilibrium geometry corresponds to the bottom of the potential well. The product potential (lower parabola) has the same shape but is shifted to a more negative potential energy by an amount ΔG, corresponding to the free energy of the reaction. The product potential is also shifted in nuclear geometry, corresponding to the rearrangement of the equilibrium nuclear geometry upon electron transfer. The reorganization energy (λ) corresponds to the energy that must be added to the reactants to move from the equilibrium reactant geometry to the equilibrium product geometry, but remaining on the reactant surface, that is without transferring an electron. In a quantized view of the simple harmonic oscillator potentials, only certain energy levels are permitted, shown as horizontal lines spaced by the quantum energy of the oscillator ħω. This energy corresponds to the characteristic frequency of the oscillator. In the quantized view, nuclear tunnelling from reactant to product depends upon the overlap of the harmonic oscillator wave functions. In this figure, we show schematically the square of the wave functions (corresponding to the probability density of the wave functions) as shaded regions. On the right, the activation energy of the reaction, corresponding to the...
Table of contents
Cover
Half Title
Title Page
Copyright Page
Table of Contents
Contributors
Abbreviations
Preface
1 Outline of theory of protein electron transfer.
2 The protein bridge between redox centres.
3 Interprotein electron transfer.
4 Computer modelling of protein–protein interactions.
5 Structure of electron transfer proteins and their complexes.
6 Photosynthetic bacterial reaction centres.
7 Copper proteins.
8 Haemoproteins.
9 Electron transfer complexes coupled to ion translocation.
10 Electron transfer reactions in chemistry. Theory and experiment.
A Electron transfer rate calculations.
B Kinetic analysis.
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Protein Electron Transfer by Dr Derek Bendall in PDF and/or ePUB format, as well as other popular books in Art & Biochemistry. We have over one million books available in our catalogue for you to explore.