![]()
IV
Evolution und Selektion
![]()
7
LUCA – letzter gemeinsamer Vorfahre allen Lebens
Armen Mulkidjanian und Dirk-Henner Lankenau
»Es ist wirklich lachhaft was so alles in den Köpfen mancher Naturforscher herumspukt, wenn sie von »Arten« sprechen. ...
Ich glaube das rührt alles daher, das Undefinierbare definieren zu wollen.«
(Charles Darwin an J.D. Hooker, 24. Dezember 1856)6
»Es ist die Gattung, nach der sich die Merkmale ergeben, und nicht die Merkmale, welche die Gattung erfordern.«
(Carolus Linnaeus, 1737)(115)
Einführung: Darwins Stammbaum des Lebens
Wie wir in den anderen Kapiteln dieses Buches sehen, versuchen viele Wissenschaftler, das Problem des Ursprungs des Lebens zu lösen. Dabei betrachten sie chemische Reaktionen, die aus einfachsten Verbindungen für Lebewesen wichtige, d.h. biologisch relevante Moleküle erzeugen können. Die Wahrscheinlichkeit, dass solche Reaktionen auch tatsächlich, plausibel auf der Urerde stattgefunden haben, wird dann im Labor experimentell überprüft. Einige Wissenschaftler wählen jedoch eine andere Herangehensweise, um den Ursprung des Lebens auf unserem Planeten zu rekonstruieren. Dieser Ansatz geht auf Charles Darwins Werk »Die Entstehung der Arten durch natürliche Zuchtwahl« zurück und beruht darauf, den Prozess der biologischen Evolution als einen Stammbaum des Lebens zu betrachten [26]. Der Baum des Lebens wurde bereits im Buch Genesis erwähnt: »Gott, der Herr, ließ aus dem Ackerboden allerlei Bäume wachsen, verlockend anzusehen und mit köstlichen Früchten, in der Mitte des Gartens aber den Baum des Lebens. ...« Darwin benutzte diese Idee des wachsenden Baumes, um die Evolution der Organismen aus einer gemeinsamen Wurzel zu veranschaulichen. Die ersten Stammbäume des Lebens, die Darwin zeichnete, waren noch nicht besonders umfangreich.
Abb. 36 Darwins drei Skizzen eines Stammbaums aus dem berühmten ersten Notizbuch B [29].
Abbildung 36 zeigt erste Skizzen des evolutionären Baums des Lebens (1837). Sie stammen aus Darwins Notizbuch B zur »Transmutation der Arten«, welches ausgestellt ist im Museum of Natural History in Manhattan, New York City [175]. Darwins Genius stellte allerdings gleichzeitig fest: »The tree of life should perhaps be called the coral of life, base of branches dead; so that passages cannot be seen.« [29]. Übersetzt bedeutet dies: »Der Baum des Lebens sollte vielleicht besser Koralle des Lebens heißen, die Wurzeln der Zweige sind (wie bei Korallen) oft abgestorben, so dass Übergänge nicht einfach erkannt werden können.« Diese Zuordnung spielt eine sehr wichtige Rolle bei unserer nun folgenden Betrachtung des gemeinsamen Vorfahren allen Lebens. In dem Buch »Die Entstehung der Arten« gab es nur eine einzige Abbildung, und diese war der nun wesentlich präzisierte Stammbaum des Lebens.
Abb. 37 Charles Darwins berühmter Stammbaum zum Ablauf von Artbildungsprozessen. Artspaltung und Neubildung sind immer begleitet von Radiation (das sind die kleinen buschähnlichen Zweige an jeweiligen Knotenpunkten) und Aussterben. Die einfachste Aufspaltung – nämlich die rein zweischenklige (dichotome) Aufspaltung in zwei Arten – ist ebenfalls nur eine sehr einfache Form der Radiation, wie ein Kreis nur eine Sonderform der Ellipse ist (Abbildung aus [26]).
Darwin verstand offenbar, dass die Zweige eines Baumes letztendlich immer aus einem gemeinsamen »Punkt« hervorgehen, wobei ihm sogar die Details der buschähnlichen Neuentstehung von Arten bewusst waren, die selbst heutzutage manchmal noch übersehen werden (Abbildung 37). Er schrieb: »Es liegt eine Erhabenheit in dieser Betrachtung des Lebens, mit seinen vielfältigen Machtebenen, eingehaucht in nur wenige (Ur)Formen oder sogar nur eine einzige.« (»There is a grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one« [28], S. 508). Zur Zeit Darwins gab es jedoch keinerlei Hinweis darauf, dass alle Lebewesen tatsächlich auf einen einzigen Vorfahren zurückgeführt werden könnten. Ein weiteres Jahrhundert musste vergehen, damit sich erwies, dass Darwin – wie so oft – vollkommen richtig lag. Heute wissen wir, dass genetische Information bei allen Organismen in DNA- und RNA-Strängen, bestehend aus Nucleinsäureketten, gespeichert ist. Darüber hinaus gibt es nur einen einzigen genetischen Code (s. unten). Demzufolge haben alle Lebewesen einen einzigen gemeinsamen Ursprung. Das lebende »Wesen«, oder besser, die lebende Entität, welche unmittelbar dem ersten Verzweigungsknotenpunkt des Stammbaumes des Lebens vorausging, wird als Progenot, Cenancestor oder LUCA (engl. Last Universal Common Ancestor oder, kontextabhängig, Last Universal Cellular Ancestor) bezeichnet. Darwins Erkenntnis der kleinen, buschähnlichen Knotenverzeigungen (Radiationen), die mit Fortschreiten der Evolution durch Aussterbeprozesse jedoch immer wieder verschwanden (Abbildung 37), führte dazu, dass wir heute auch von LUCAS sprechen (engl. Last Universal Common Ancetral State) [93] – also einem nicht-entitären Urzustand, wenn Details des Ursprungs um den ersten Knotenpunkt erörtert und erforscht werden sollen7.
Crashkurs in Biologie
Bevor wir die Eigenschaften von LUCA beschreiben, scheint es uns sinnvoll, die Grundlagen der Biologie vorzustellen. Es ist die Geschichte der Zelle, ihrer Teilung und Vermehrung. Eine Chronologie dieser Geschichte ist in Anhang A dargestellt.
Der Aufbau der Zelle
Weil alle lebenden Organismen zellulär sind, war LUCA irgendwann auch zellulär organisiert und darum interessieren uns alle Details moderner Zellen. Heute kennen wir den Aufbau und die molekularen Lebensfunktionen bis auf die atomare Ebene genau. Jede Zelle ist von einer vollständigen Membranhülle umgeben. Den Membranen aller Lebewesen sind zwei Merkmale gemeinsam, nämlich erstens eine Lipiddoppelschicht und zweitens Membranproteine, die die Doppelschicht durchstoßen.
Lipide bilden die flüssige Grundstruktur aller biologischen Membranen. Sie bestehen aus wasserabstoßenden Kohlenstoffketten und einer geladenen, polaren, d.h. hydrophilen8 Kopfgruppe. In Lipid-doppelschichten sind die hydrophoben9 Schwänze stets einander zugewandt, während die hydrophilen Köpfe nach außen und innen zeigen und an das umgebende wässrige Medium grenzen. Durch die Abgrenzung durch Membranen erzeugt die Zelle eine innere Umwelt, die in ihrer Zusammensetzung völlig anders ist als die Außenwelt. Damit hebt sich jede Zelle als konkrete, lebende Entität von oft lebensfeindlichen Umwelten ab.
Hinsichtlich der Zellstruktur unterscheiden sich grundlegend zwei Gruppen von Organismen, nämlich Prokaryonten und Eukaryonten10. Bei Eukaryonten ist die gesamte Erbinformation, die DNA, von einer Doppelmembran umgeben. Diese Struktur ist die von Robert Brown (siehe Anhang A) 1831 Areola genannte, lichtbrechende Struktur, der Zellkern (Karyon bedeutet im Griechischen Kern). Prokaryonten besitzen ein Nucleoid, in dem die DNA »lagert«. Das Nucleoid ist jedoch nicht von einer Doppelmembran umschlossen.
Außerhalb des Kerns besitzen Eukaryontenzellen im Zellplasma membranumhüllte Kompartimente und Strukturen. Diese sind insbesondere die Chloroplasten, Mitochondrien, Membransysteme wie der Golgiapparat und das endoplasmatische Reticulum (ER) sowie ein Zellskelett. Mitochondrien und Chloroplasten waren ursprünglich eigenständige Prokaryonten, die sehr früh im Laufe der Evolution in einen Vorläufer aus der Stammgruppe der Eukaryonten integriert wurden. Seit dieser Vorzeit erfüllen die symbiotischen Organellen lebensnotwendige Funktionen in jeder Eukaryontenzelle. In den Mitochondrien findet die Energiegewinnung im Citratcyclus und der Atmungskette statt. Sowohl Pflanzen als auch Tiere besitzen Mitochondrien. Chloroplasten kommen bei Pflanzen und Algen vor. In ihnen findet die Photosynthese statt. Bei der Photosynthese wird Lichtenergie dazu benutzt, Elektronen von anorganischen Stoffen abzuziehen und dann mit Hilfe von Enzymen auf Kohlenstoffdioxid (CO2) zu übertragen. Dadurch entstehen verschiedene Verbindungen, die zur Ernährung dienen können.
Die genetische Information, die innerhalb der DNA- und RNA-Stränge gespeichert vorliegt, kann man vergleichen mit dem Barcode (Strichcode) auf den Waren eines Supermarktes. Die DNA- und RNA-Stränge stellt man sich dabei am besten als Drachenschnur vor, auf der dieser Barcode gespeichert ist. In der DNA gibt es jedoch nur vier »Striche«, die Nucleotide A (Adenin), C (Cytosin), G (Guanin) und T (Thymin). Sie sind durch Zucker-Phosphat-Bindungsglieder aneinander gekoppelt. A und T einerseits sowie C und G andererseits können Paare bilden, dadurch können zwei DNA-Moleküle miteinander interagieren und doppel-helikale Strangstrukturen bilden.
Sowohl bei Prokaryonten als auch bei Eukaryonten wird die genetische Information in der DNA durch Replikation des Genoms (Verdopplung der »Drachenschnur«) erhalten [178, 179]. Die Replikationsreaktion wird so durch das DNA-Polymerase-Enzym durchgeführt [98], sodass jede Zelle eine exakte Kopie des Genoms11 der elterlichen Zelle erhält. Die genetische Information in der DNA (Barcode) wird durch Transkription der Gene in einzelne mRNA-Stränge (messenger-RNAs, Boten-RNAs) überführt und für weitere zellphysiologische Prozesse zur Verfügung gestellt. Bei allen Organismen wird die Transkription, d.h. die Synthese von RNA-Polymer entlang des DNAPolymers, durch das Enzym RNA-Polymerase erledigt [100].
Wie DNA besteht jede mRNA aus Ribonucleotid-Bausteinen mit je einer von vier möglichen Basen: A (Adenin), C (Cytosin), G (Guanin) und U (Uracil). Das bedeutet, dass ein RNA-Strang anstelle eines T stets ein U enthält. Die mRNAs werden dann in die verschiedenen Aminosäure-Sequenzen all der Proteine eines Organismus übersetzt (translatiert). Jedes Aminosäure-Kettenglied wird dabei durch ein oder mehrere Basentripletts (Codons) codiert, z.B. UUA oder UUG für die Aminosäure Leucin. Auch Signale für Start (AUG) und Ende der Translation (UAG, UAA oder UGA) sind in dem Code enthalten [134, 162]. Dieser Code gilt universell für alle Lebewesen und Viren.
Die Translation der genetischen Information in der mRNA erfolgt im Ribosom. Die Ribosomen selbst sind aus RNA und Proteinen zusammengesetzte Nano-Maschinen, die die Übersetzung des genetischen Codes in eine bestimmte Reihenfolge von Aminosäurekettengliedern gewährleisten. Die Proteinsynthese im Ribosom kann sowohl frei im Cytoplasma als auch an den Zellmembranen stattfinden.
Die mechanisch-molekulare Verbindung von mRNA-Sequenz zu Proteinsequenz wird von der Zelle durch transfer-RNA (tRNA) realisiert. Bakterienzellen haben etwa 50 verschiedene Sorten von tRNA-Molekülen, wobei jede etwa 75 Nucleotide lang ist. An ihr CCA-3′-Ende wird über eine Esterbindung jeweils eine bestimmte Aminosäure gebunden. In den 75 Nucleotiden der tRNA befindet sich eine drei Nucleotide umfassende Sequenz, die Anticodon genannt wird. Das Anticodon bindet im Ribosom (wie ein Magnet) an sein komplementäres Gegenstück in der mRNA und positioniert seine charakteristische Aminosäure im Ribosom; die Aminosäuren werden zuvor durch ein tRNA-spezifisches Enzym an ihre tRNA geknüpft. Diese Komplexe heißen Aminoacyl-tRNAs. Wenn im Ribosom zwei benachbarte Aminoacyl-tRNAs ihre jeweiligen Aminosäuren benachbart positioniert haben, werden die beiden Aminosäuren in einer Peptidbindung zu einer Dipeptidsequenz, zwei Kettengliedern eines werdenden Proteins, verknüpft. Dies ist die Peptidyltransferase-Reaktion. Eine Rückcodierung (reverse Transkription) von RNA zu Doppelstrang-DNA ist möglich und ursprünglich [20, 170], während es eine umgekehrte Übersetzung (reverse Translation) vom Protein zur RNA nicht gibt. Dies ist das zentrale Dogma der Molekularbiologie [23, 23a].
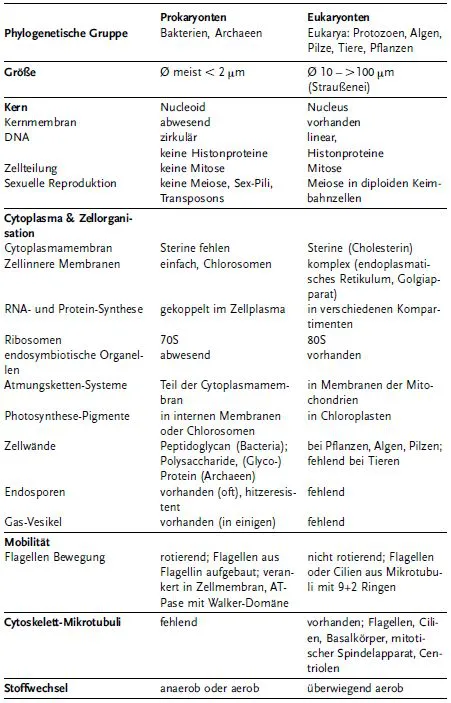
Prokaryontenzellen und Eukaryontenzellen unterscheiden sich in vielen Details und nur wenige Eigenschaften, Prozesse und Gene sind identisch. Tabelle 1 zeigt einen klassischen Vergleich der Eigenschaften dieser zwei großen Gruppen.
Wie bestimmt man die Verwandtschaft zwischen Organismen?
Während Chemiker im 20. Jahrhundert die Bausteine des Lebens entdeckten und beschrieben, durchdachten und erforschten Biologen die körperinternen und die äußeren wechselseitigen Beziehungen der Lebewesen. Auch die Wechselwirkungen der Organismen in Ökosystemen wurden wichtig. Wieder einmal war Darwin der Pionier, der als Erster das »Denken in Populationen« begriffen hatte [114].
Tabelle 1 Vergleich der Zellen von Prokayonten und Eukaryonten.
Essenziell für die Populationsbiologie und heutige Ökologie war seit jeher ein kausal-exaktes Verständnis der biologischen Vielfalt auf der Erde. Phänomenologie der Natur war und ist ein seriöses Anliegen. Der dazugehörige Wissenschaftszweig heißt Systematik. Wie benenne ich ein Tier? Wie heißt eine Pflanze? Was ist eine Flechte, ein Ameisenstaat, ein Bakterium und was ist ein Virus? Hier herrschen oft konfuse Vorstellungen. Wissenschaftliche Namen haben die Aufgabe, ein lebendes Individuum eindeutig zu bezeichnen. Die einfache wissenschaftliche Benennung von Organismen ist die »Nomenklatur«. Carl von Linné publizierte ab 1753 sein Werk »Systema Naturae«, in dem er die bis heute gebräuchliche binome Nomenklatur der Arten einführte. Einer Gattung (lat. genus) wurden ein oder mehrere Arten (lat. species) zugeordnet – z. B. Homo (genus) sapiens (species). Bis heute bestimmt man Arten in der Natur pragmatisch mit Bestimmungsschlüsseln, in denen sogenannte diagnostische Merkmale (z. B. bei Pflanzen kreuzgegenständige Blätter, Anzahl der Staubgefäße, Farbe der Blüten, bei Tieren die Anzahl der Wirbel, relative Größe und Anzahl der Fühlerglieder usw.) als Kriterien der Unterscheidung herangezogen werden. Der binäre Artbegriff funktioniert allerdings nur exakt bei sich sexuell vermehrenden vielzelligen Organismenpopulationen (Metazoen). Dies hängt mit einem Evolutionsgesetz zusammen, welches Mullers Ratsche genannt wird und für die überwältigende Mehrheit von Tier und Pflanzenpopulationen gilt [102]. Auch für Bakterien, z.B. bei Escherichia coli, wird die binäre Nomenklatur benutzt, sie ist jedoch nicht vergleichbar mit der Nomenklatur sexueller Metazoen. Einzellige Mikroorganismen stellen Urformen von relativ selbstständigen Keimbahnen dar, die hohe Reproduktionszahlen aufweisen und vielfältig mit ihrem Lebensraum interagieren.
Ein bedeutender Wechsel vollzog sich in der biologischen Systematik 1936, als die Taxonomie von Carl von Linné zur phylogenetischen Systematik oder Kladistik von Willi Hennig wechselte [71, 152, 153]. Der erste Schritt in Hennigs Methode war die strikte Beschränkung auf abstammungsmäßige, d. h. phylogenetische oder genealogische Verwandtschaft. Mit dieser Methode wurden nicht mehr ausschließlich die morphologischen Eigenschaften zur Verwandtschaftsaufklärung herangezogen, sondern Hennig bemühte sich, ähnlich Poppers (1935) deduktivem Ansatz [177], ganzheitlich alle Kriterien der Verwandtschaft zu berücksichtigen und immer wieder neu zu bewerten. Der Begriff phylogenetische Systematik existierte bereits vor Hennigs Ausarbeitung. Es war auch schon lange akzeptiert, dass nur Abstammungsähnlichkeiten (echte Homologien, bei Genen heißen sie Orthologien) phylogenetische Verwandtschaft anzeigen. Es gab jedoch weder akzeptierte Kriterien zur Ermittlung von Homologien noch eine wissenschaftlich exakte Methode zur Rekonstruktion phylogenetischer Beziehungen. Die neue Terminologie der phylogenetischen Systematik entwickelte Hennig während der Kriegsjahre 1939–1945. Er nannte die alten Merkmale »plesiomorph« und die neu entstandenen Eigenschaften »apomorph«. Bei der Ermittlung der nächsten Verwandtschaft zwischen zwei Arten sind nicht die plesiomorphen, sondern die gemeinsamen, relativ (gegenüber entfernter verwandten Arten) neuen, apomorphen Merkmale entscheidend [72, 152]. 1950 stellt Hennig dann den Bezug zum Begriff »monophyletisch« her [70], der zurückgeht auf den von Charles Darwin verehrten Verfechter des Evolutionsgedankens in Deutschland, Ernst Haeckel [62]. Monophyletische Gruppen sind Taxa, deren Mitglieder alle von einem unmittelbaren, gemeinsamen Vorfahren abstammen. Gruppen, die man fälschlicherweise aufgestellt hat, werden polyphyletisch genannt, wenn Organismen mit verschiedenen Vorfahren zusammengruppiert werden und paraphyletisch, wenn nur eine Teilmenge der tatsächlich verwandten Arten berücksichtigt wurde. Eine Abfolge der zugrunde liegenden Artspaltungsereignisse – also Darwins »origin of species« – wurde von dem deutschen Neodarwinisten Bernhard Rensch als Kladogenese (Stammverzweigung) bezeichnet [144]. Ernst Mayr, in Berlin ein Schüler von Bernhard Rensch, nannte Hennigs Methode dann »kladistisch« [11, 115]. Die moderne molekulare Systematik basiert auf dem genetischen Aspekt der Definition des Lebens12 durch Hermann Muller: Es sind solche Entitäten als lebend zu definieren, die die Fähigkeit der Vermehrung, der Variation und der Vererbung besitzen [129]. Demnach sind in einem modernen System biologischer Organismen nicht jene miteinander verwandt, die sich im Aussehen am meisten ähneln, sondern jene, die gemeinsam ähnliche Gene besitzen und evolutionär miteinander verwandt sind. Damit ist das heutige System der Organismen gewissermaßen eine Reflexion von Charles Darwins erstem Stammbaum des Lebens (siehe Abbildung 36). In einem Brief an Thomas Huxley sah Darwin die Zeit voraus, »when we shall have very fairly true genealogical trees of each kingdom of nature« [27].
Die drei Domänen des Lebens
Ein anderer Paradigmenwechsel ereignete sich in der Mikrobiolo...