Keith T. Sillar, Laurence D. Picton, William J. Heitler
This is a test
This is a test
Buch teilen
English
ePUB (handyfreundlich)
Über iOS und Android verfügbar
eBook - ePub
The Neuroethology of Predation and Escape
Keith T. Sillar, Laurence D. Picton, William J. Heitler
Angaben zum Buch
Buchvorschau
Inhaltsverzeichnis
Quellenangaben
Über dieses Buch
THE NEUROETHOLOGY OF PREDATION AND ESCAPE
To eat and not get eaten is key to animal survival, and the arms race between predators and prey has driven the evolution of many rapid and spectacular behaviours.
This book explores the neural mechanisms controlling predation and escape, where specialisations in afferent pathways, central circuits, motor control and biomechanics can be traced through to natural animal behaviour.
Each chapter provides an integrated and comparative review of case studies in neuroethology. Ranging from the classic studies on bat biosonar and insect counter-measures, through to fish-eating snails armed with powerful neurotoxins, the book covers a diverse and fascinating range of adaptations. Common principles of biological design and organization are highlighted throughout the text.
The book is aimed at several audiences:
for lecturers and students. This synthesis will help to underpin the curriculum in neuroscience and behavioural biology, especially for courses focusing on neuroethology
for postgraduate students. The sections devoted to your area of specialism will give a flying start to your research reading, while the other chapters offer breadth and insights from comparative studies
for academic researchers. The book will provide a valuable resource and an enjoyable read
Above all, we hope this book will inspire the next generation of neuroethologists.
Häufig gestellte Fragen
Wie kann ich mein Abo kündigen?
Gehe einfach zum Kontobereich in den Einstellungen und klicke auf „Abo kündigen“ – ganz einfach. Nachdem du gekündigt hast, bleibt deine Mitgliedschaft für den verbleibenden Abozeitraum, den du bereits bezahlt hast, aktiv. Mehr Informationen hier.
(Wie) Kann ich Bücher herunterladen?
Derzeit stehen all unsere auf Mobilgeräte reagierenden ePub-Bücher zum Download über die App zur Verfügung. Die meisten unserer PDFs stehen ebenfalls zum Download bereit; wir arbeiten daran, auch die übrigen PDFs zum Download anzubieten, bei denen dies aktuell noch nicht möglich ist. Weitere Informationen hier.
Welcher Unterschied besteht bei den Preisen zwischen den Aboplänen?
Mit beiden Aboplänen erhältst du vollen Zugang zur Bibliothek und allen Funktionen von Perlego. Die einzigen Unterschiede bestehen im Preis und dem Abozeitraum: Mit dem Jahresabo sparst du auf 12 Monate gerechnet im Vergleich zum Monatsabo rund 30 %.
Was ist Perlego?
Wir sind ein Online-Abodienst für Lehrbücher, bei dem du für weniger als den Preis eines einzelnen Buches pro Monat Zugang zu einer ganzen Online-Bibliothek erhältst. Mit über 1 Million Büchern zu über 1.000 verschiedenen Themen haben wir bestimmt alles, was du brauchst! Weitere Informationen hier.
Unterstützt Perlego Text-zu-Sprache?
Achte auf das Symbol zum Vorlesen in deinem nächsten Buch, um zu sehen, ob du es dir auch anhören kannst. Bei diesem Tool wird dir Text laut vorgelesen, wobei der Text beim Vorlesen auch grafisch hervorgehoben wird. Du kannst das Vorlesen jederzeit anhalten, beschleunigen und verlangsamen. Weitere Informationen hier.
Ist The Neuroethology of Predation and Escape als Online-PDF/ePub verfügbar?
Ja, du hast Zugang zu The Neuroethology of Predation and Escape von Keith T. Sillar, Laurence D. Picton, William J. Heitler im PDF- und/oder ePub-Format sowie zu anderen beliebten Büchern aus Biowissenschaften & Zoologie. Aus unserem Katalog stehen dir über 1 Million Bücher zur Verfügung.
Very few species of animal are active only in complete darkness. At any moment in time, half of our planet is bathed in light radiating from the sun, while the other half remains partially lit by reflections from the moon, distant starlight, bioluminescence and human light pollution. It is thus unsurprising that many animals make use of some form of vision to provide information about the environment. Vision often provides the earliest cues for the detection of a predator or prey; it usually has a longer range than hearing, and is more precise and immediate than smell. Even when other sensory modalities do take the lead, vision is almost always additionally involved in the pursuit of prey, the guidance of a predatory strike or the coordination of an escape manoeuvre.
The information provided by vision depends on the complexity and sophistication of the eye, but even the simplest eyes are adept at detecting changes in light intensity, and these temporal cues are often important signals warning of the approach of danger. Many animals, particularly sessile forms such as barnacles, tubeworms or bivalve molluscs, display a shadow withdrawal reflex in response to a sudden dimming of the light. In these cases, the image detail is irrelevant, and it is the overall level of environmental illumination that matters. However, for many other animals, the image is crucial, because it enables the eye’s owner to determine where the object of interest is located. Indeed, some authorities argue that it was the evolution of image-forming eyes, and a consequent step increase in the speed of the predator-prey arms race, that led to the massive proliferation in animal diversity about 540 million years ago, known as the Cambrian Explosion (Parker, 1998).
In this chapter, we will cover some of the key case studies in neuroethology that have informed our understanding of how animals use vision to detect, identify and respond to predators and prey. We first outline important principles of visual processing, including properties of light and the concepts of foveae, acuity and sensitivity. We then look at how meaningful features of an image are extracted as information passes through the visual pathway, in order to identify objects as predator or prey, using the toad as a model system. In the next part of the chapter, we outline the neurobiology of infrared vision in snakes. These ambush predators possess a highly specialised infrared visual system used to detect and target their warm-blooded prey. Finally, we will look at specialisations of the visual system in aerial predators that enable them to capture prey while simultaneously coordinating flight, using dragonfly vision as an exquisite example.
1.1 The electromagnetic spectrum
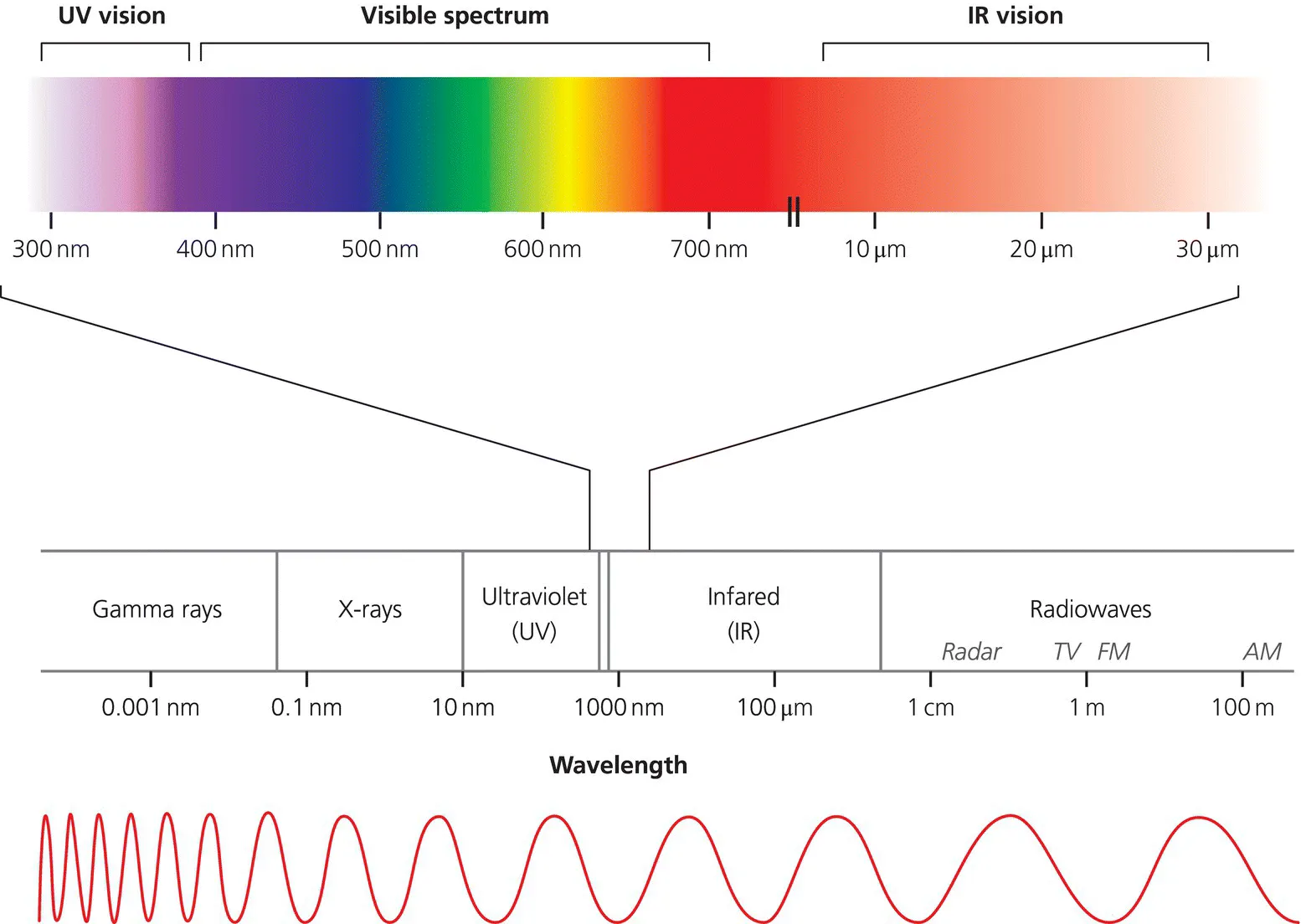
Light is a form of electromagnetic radiation (Figure 1.1) that consists of waves of energy that radiate through the air1 at roughly 300 000 km/s. The intensity of light is determined by the amplitude of these waves, while what we perceive as colour is encoded in their wavelength.
Figure 1.1 The electromagnetic (EM) spectrum. EM radiation spans a range of wavelengths, from low-energy, long wavelength radio waves to high-energy, short wavelength gamma rays. Most animals detect light only within a very narrow range of wavelengths; for example, the ‘visible spectrum’ in humans is between is about 390–700 nm. Some animals are also capable of detecting ultraviolet radiation (insects, birds and fish) and infrared radiation (crotalid and boid snakes, fire-seeking beetles and vampire bats).
For many animals, including humans, visible light spans a range of wavelengths between approximately 390 nm and 700 nm (the visible spectrum; Figure 1.1). We perceive these wavelengths as the colours of the rainbow, and we are able to do so because we possess a trichromatic visual pigment system. The cone cells of the human retina contain photo-pigments that absorb either red, green or blue wavelengths of light that has bounced off objects2 in the environment, and our brains then interpret the mixtures of cone signals as different colours. However, cones have a relatively high response threshold, and thus only respond if the light is quite bright. We possess an additional set of photoreceptors, the rod cells, which absorb light across the full visible spectrum and, thus, are unable to distinguish between different colours, but are very sensitive to low light levels. The rod system therefore only mediates greyscale imaging (and the rod pigments are completely bleached in bright daylight), but it can function at low ambient light levels, in which the cone system is effectively blind. The two systems overlap and operate in parallel at intermediate light levels, such as twilight. Not all animals possess colour vision but, for those that do, it can be very useful in detecting warning coloration, such as aposematic beetles that warn predators they are distasteful.
Many animals are able to detect wavelengths of light beyond the spectrum to which humans are restricted (Figure 1.1). For example, many insects, birds and fish can visualise shorter wavelengths of light in the ultraviolet (UV) range, and they do so to detect a range of important ecological features, from enticing landing strips on flowers (bees) to the detection of aerial insect prey against the sky (dragonflies)3. One of the few mammals that can see in UV is the arctic reindeer (caribou), and here the key advantage seems to be contrast enhancement (Hogg et al., 2011). The ground is often snow-covered, and predators such as wolves, with white fur, are hard to detect in the normal visible spectrum. However, snow is a strong reflector of UV, while fur is a strong absorber, which means that, in the UV spectrum, the wolf stands out clearly against a snowy background.
At the other end of the spectrum, some animals can also detect light in the longer wavelength infrared (IR) range (see Campbell et al. (2002) for a general review). Fire-seeking beetles, for instance, can detect IR light emitted by forest fires at a distance of up to 80 km, allowing them to navigate to freshly burnt wood, into which they lay their eggs (Schmitz and Bleckmann, 1998). Vampire bats possess IR-detecting receptors on their facial nose-leaf that are used to locate the most nutritious, blood-rich regions of a target prey animal. However, the most sophisticated IR visual system belongs to crotalid and boid snakes, which includes pit vipers, boas and pythons. These snakes possess highly specialised IR-detecting organs on their snout which allow them to detect, locate and capture their warm-blooded prey, even in what we would consider to be the dark (see Section 1.5).
Many animals, including a number of arthropods and cephalopods, are also highly sensitive to another property of light: polarisation. Mantis shrimp are the most sensitive – they have 12 visual pigments, and not only can they detect UV and IR radiation, but also circularly and linearly polarised light! Polarisation sensitivity is probably mainly used for navigation, but it can also be used for detecting certain types of prey – for instance, squid use polarized light sensitivity to visualise otherwise invisible, transparent zooplankton (Shashar et al., 1998).
1.2 Eyes: acuity and sensitivity
The quality of an eye and, therefore, its usefulness to either predator or prey, strongly depends on two key factors:
Sensitivity – the amount of light energy that it can capture from a given source.
Resolution (acuity) – the accuracy with which it can determine the spatial origin of that source.
Sensitivity depends partly on receptor physiology, but also on eye str...