![]()
PART I
Foundations
‘Science is, I believe, nothing but trained and organised common sense’.
Thomas Henry Huxley (‘Darwin’s bulldog’) Collected Essays, vol. 3, 1893
The exciting challenge that this book sets itself is to explain to you what is known about the ways in which the development of animals and plants evolves; and also, in the final chapter, to explain what is not yet known, and thus what are the key questions for future research. We begin to approach this challenge on the foundations provided by the first four chapters, as follows:
Chapter 1: We start by examining the reason why understanding egg-to-adult development is absolutely crucial for a complete picture of the evolution of multicellular creatures.
Chapter 2: This chapter deals with the forerunners of present-day evolutionary developmental biology (or evo-devo), the way in which evo-devo has arisen, and the nature of evo-devo today, including the range of approaches that can be included within it.
Chapter 3: Here, the focus is on the nature of the developmental process, in terms of the things cells do, which are the results of an interplay between genes, their products, an array of other molecules and, often, environmental effects.
Chapter 4: Our focus now shifts to the structure of natural populations, since it is in these that all evolution happens. We examine their spatial, age and genetic structure. We then look at ways in which the genetic structure of populations can be altered by natural selection.
![]()
1.1 From Darwin to Development
1.2 Development; and Evolutionary Changes in Development
1.3 Development and the Realm of Multicellularity
1.1 From Darwin to Development
The theory of evolution, established by Charles Darwin more than 150 years ago, and still itself evolving, is one of the most impressive products of science. As Darwin said1 in the closing paragraph of The Origin of Species: ‘There is grandeur in this view of life’ – a view in which many diverse creatures, both past and present (Fig. 1.1), have been brought into existence by natural processes, and in particular by the interplay between two such processes – heritable variation and natural selection.
Darwin marshalled a wide range of evidence in support of his theory. He drew on information from animal and plant breeding, fossils, behaviour, morphology, embryology and geography, among others. And he used all of these to build a sound basis for his key contribution to evolutionary theory: natural selection.
But evolution is, as noted above, the result of an interplay between two things – heritable variation and natural selection – it is not explicable by either of these on its own. The fact that Darwin was unable to enlighten us as much about the former as about the latter was hardly due to an oversight on his part; rather, it was due to limitations on what was generally known at the time in this area of ‘heritable variation’.
It is worth dissecting this phrase, because it includes both the inheritance of genetic variation and the process of development through which phenotypic variation is produced. Darwin was aware of the problem that there was not, in the 1850s, a clear understanding of how inheritance worked, but proceeded as best he could regardless. He later tried to supply a theory of inheritance – ‘pangenesis’ – but got it wrong. He was doubtless also aware that there was not a clear understanding of how egg-to-adult development worked, in terms of causal mechanisms, but he proceeded to use the information on descriptive and comparative embryology that did exist in the 1850s to good effect. As he remarked (Chapter 13), ‘community in embryonic structure reveals community of descent.’
The two most important things that have happened since Darwin’s synthesis of the evidence for evolution in 1859 have been the incorporation of genetics and developmental biology into the ‘big picture’, with the result that it has even more grandeur than before. The incorporation of genetics, which came first (Appendix 1), had both positive and negative effects on the incorporation of developmental biology that followed, is still in full swing, and is the subject of this book.
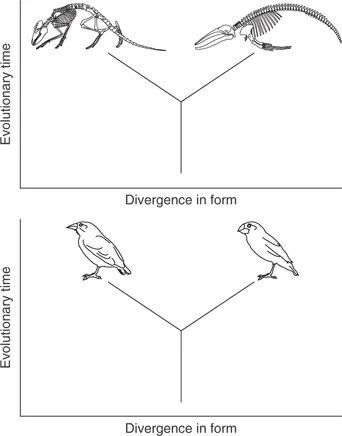
But why, actually, is development so important for evolutionary theory? There is a very specific and compelling answer to this question. It relates to the ways in which evolution can and cannot produce one type of animal or plant from another. This point is best made in relation to the type of evolutionary trees typically found in papers and books on the subject, two of which are shown in Fig. 1.2. Notice that in both trees the vertical axis is some measure of time, while the horizontal axis is some measure of difference in the morphology of the animal concerned, in one case ‘generalised’ and hard to quantify, in the other case a very specific measure of a particular structure (the depth of a bird’s beak).
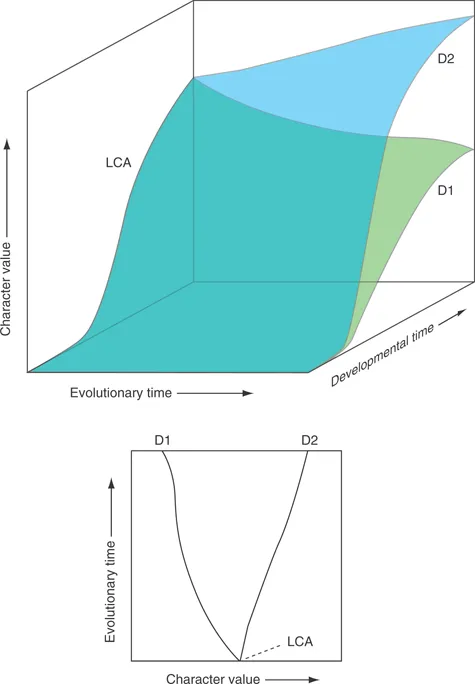
In a very important sense, both of these trees represent impossible evolutionary transitions. They both employ the familiar shorthand method of representing an animal by one particular stage of its life-cycle – the adult. But evolution cannot make one kind of adult directly from another. Rather, it can only make a new kind of adult by altering, over a period of generations, the egg-to-adult developmental trajectory (Fig. 1.3).
So, an adequate theory of evolution must include not only an account of how fitness differences cause changes at the population level, but also an account of how the developmental differences that natural selection acts upon arise in the first place. And these latter differences cannot simply be written off as mutations, because a mutation is merely a change in the DNA sequence of a gene. If the gene that mutates causes the developmental trajectory to alter, then we need to know how this happens. Furthermore, developmental trajectories can in most cases be influenced by environmental factors as well as by genes. This is true not just of extreme cases, such as the production of male or female forms in turtles by egg incubation temperature, but also of more subtle cases, such as slight differences in the amount of left-right body asymmetry (often referred to as fluctuating asymmetry) that can be the result of variation in temperature and other environmental factors.
Such considerations give the environment not just one role in evolution – that of selective ‘sieve’ – but rather two, with the other being a role in the production of the variation in the first place. Of course, non-heritable variation, or phenotypic plasticity, cannot itself contribute to evolution, precisely because of its non-heritable nature. But if different genotypes differ in their pattern of developmental response (or their developmental reaction norm) to environmental variation, as is now widely known to occur, then this provides material for evolutionary change. Indeed, all evolutionary theory that deals with phenotypes that are completely genetically determined can be regarded as a subset of more general evolutionary theory in which the determination of developmental trajectories, and hence of phenotypes, is more complex.
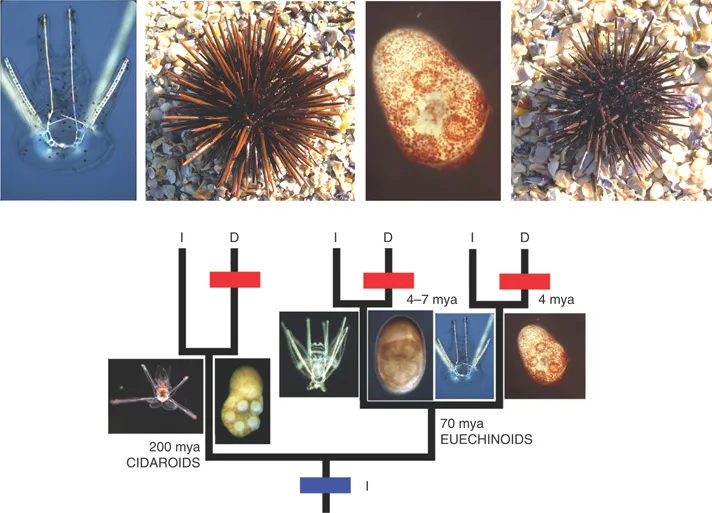
What evolve, therefore, are not just adult animals or plants, but rather complete life-cycles. Furthermore, we should not think of pre-adult stages as evolving ‘in order to enable adult forms to evolve’. This overly ‘adultocentric’ view of things (as recently criticised by the Italian biologist Alessandro Minelli2) is misleading. Instead, what happens is that there are variations at all developmental stages. In each case, some variants may be fitter than others, either because of the advantages they possess at that stage or because of advantages that accrue further downstream in the developmental pathway or, of course, for both reasons. In some cases, particularly in animals with complex life-cycles (or ‘indirect development’ – i.e. development to adult via a larval form), evolution of larval stages may occur quasi-independently of evolution of the adult. This is true, for example, in the case of evolutionary switches between plankton feeding and yolk feeding in echinoderm larvae3, where the plankton-feeders have ‘arms’ that the yolk-feeders do not (Fig. 1.4), but this does not lead to a corresponding difference in the adults.
Having now seen that the case for the centrality of development in the evolutionary process is unassailable (but with a caveat to be discussed in Section 1.3), we need to examine development itself, and also to ask about the ways in which it can evolve. This approach (Section 1.2) will reveal several problems, some of which can be easily remedied at our current stage of knowledge, but some of which cannot. These problems include: the absence of some key terms; the previous over-emphasising of some processes (e.g. heterochrony); the need to connect organism-level observations with both molecular and populational ones; and the crucial issue of whether development in some sense guides evolution. This last issue is perhaps the most fascinating of all but is also the hardest to deal with and the most controversial. At stake here is the question of whether the structure of the developmental variation available to natural selection can influence the direction that evolution takes, rather than this direction being entirely set by selection alone; and if the former, then whether the role of development is merely negative (‘constraint’) or both positive and negative (‘bias’). This will be discussed in detail in Chapter 13.
1.2 Development; and Evolutionary Changes in Development
The development of any animal or plant can be thought of as a time-sequence of more or less well-defined stages. The simplest kind o...