This chapter reviews the carbohydrate physiology and biochemistry of Allium species. We limit discussion mainly to issues peculiar to these species, and particular reference will be made to soluble carbohydrates, their distribution, and the synthesis of fructans. Much of our comment refers to work with onion (A. cepa). Some reference is made to investigations of other Allium species, but this is limited, since knowledge of interspecies differences is limited. No reference is made to the broader issues of carbohydrate metabolism, such as sucrose synthesis, which are beyond the scope of this chapter and for which readers are referred to other sources.1, 2, 3
II. COMPOUNDS AND THEIR DISTRIBUTION
The nonstructural carbohydrates in Allium spp. include glucose, fructose, and sucrose together with a series of oligosaccharides, the fructans.4, 5, 6, 7, 8, 9, 10 Fructans occur in the Asteraceae,11 but more widely in the monocotyledons, especially in the Amaryllidaceae, Iridaceae, Liliaceae, and Poaceae.12 In all species fructans are considered to be fructose polymers containing a single glucose residue. Early reports12-13 suggested only fructose was present, but later Palmer14 demonstrated the presence of glucose in fructans from a number of sources. The mechanism of fructan synthesis, demonstrated by Edelman and Jefford15 supports the presence of a glucose residue in fructans because fructose is transferred from one sucrose molecule to another sucrose molecule.
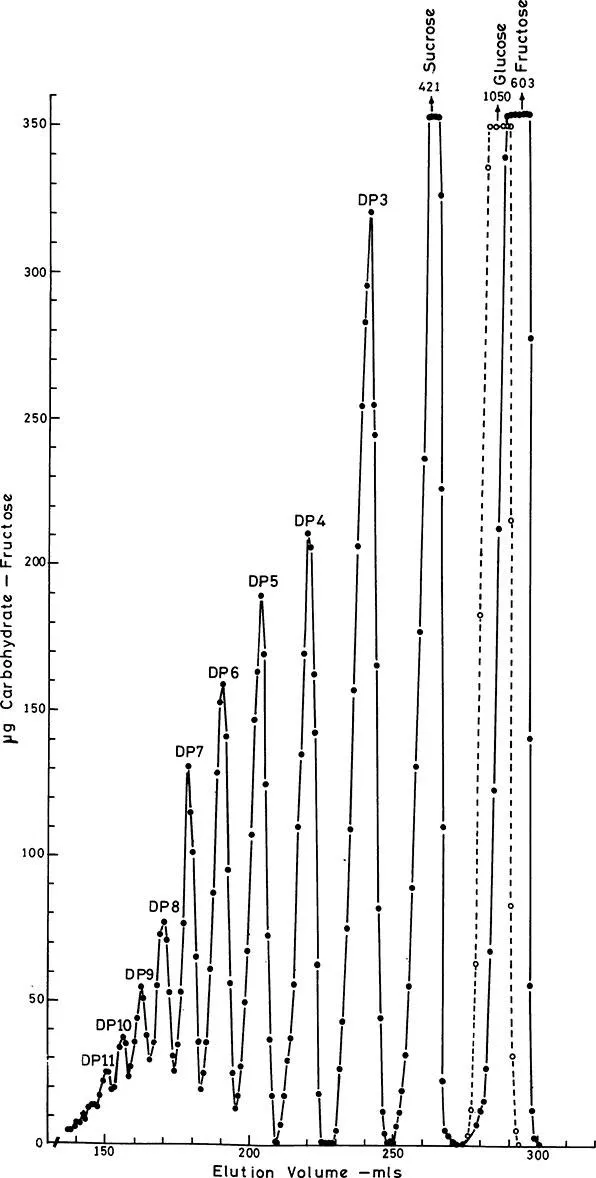
It is important to appreciate that in plants, three different trisaccharides are potentially possible, and all three have been identified in a variety of plants.5,16,17 They are 1F-fructosylsucrose, 6F-fructosylsucrose, and 6G-fructosylsucrose. These three trisaccharides arise as a result of the transfer of a fructosyl residue from one sucrose molecule to one of three primary hydroxyl groups of another sucrose molecule. The products of such a reaction are one of three possible trisaccharides and glucose. For a simple, diagrammatic representation of the structure of these trisaccharides readers are referred to Henry.43 In onion the trisaccharides 1F-fructosylsucrose and 6G-fructosylsucrose have been identified,5,9,18 and both have been reported in leek.5 These isomers have different functions in fructan synthesis (see Section IV). The addition of further fructosyl groups to these trisaccharides leads to the production of tetrasaccharides and higher polymers, giving rise to many possible polymers. The degree of polymerization (DP) of fructans can be quite different in different species. Polymers up to a DP of 260 have been reported in grasses.19 Figure 1 shows the separation of polymers from onion.
Starch is usually not detectable in onion, but Wilson et al.20 have reported starch inclusions in the chloroplasts of the uniseriate layer of cells around the vascular bundles in onion leaves. Starch was not found in the chloroplasts of the other mesophyll cells. The plants used in their study were 10 weeks old, but it is not clear whether they were grown in bulb-inducing conditions or not. When there is a substantial translocation of carbohydrate to the leaf base, starch may not be present even in the vascular sheath cells.
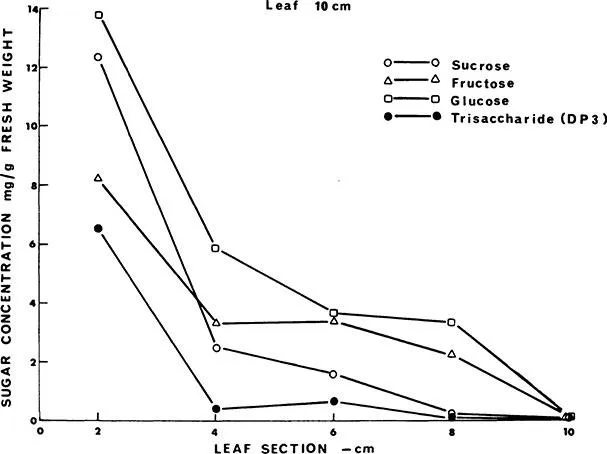
The soluble carbohydrate concentration per unit of fresh weight of onion leaf is lowest at the top and highest in the basal part (Figure 2). This pattern does not vary substantially with leaf size or level of insertion. Sugar concentration increases in leaf blades, pseudostem, and bulbs with both increasing age and light intensity7 (Figure 3).
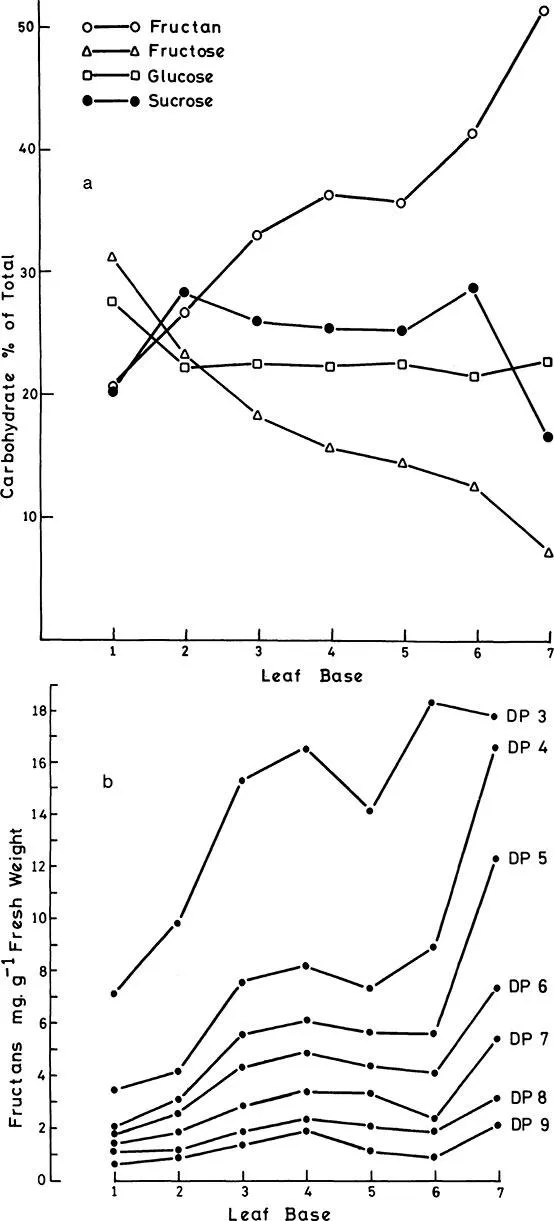
Bacon5 examined the distribution of the lower molecular weight fructans (up to DP 5) in onion bulbs and found these compounds to be absent from outer, older leaf bases and present in increasing amounts from the outer to the inner leaf bases. De Miniac8 reported a similar distribution. These observations were extended by Darby shire and Henry9 who reported the distribution of fructans of DP 3 to DP 9 (Figures 4a and b). Free fructose concentration was highest in the outer leaf bases and lowest in the innermost, an observation made also by Rutherford and Whittle.9a Glucose and sucrose did not change across the bulb.
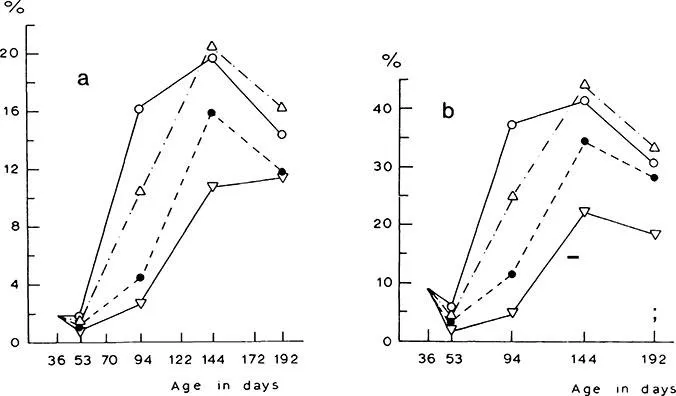
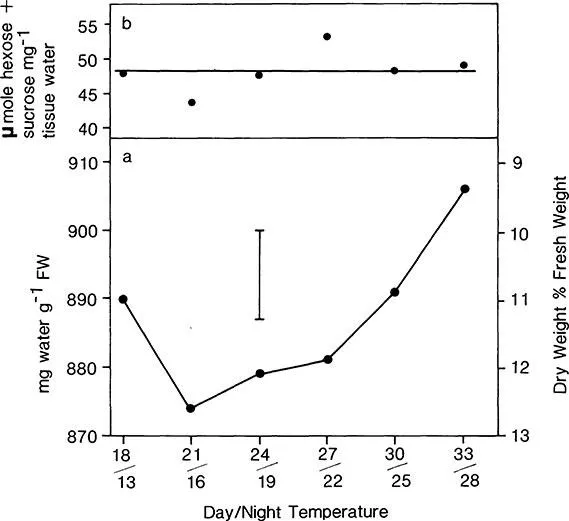
Under controlled conditions Steer21 examined the influence of day/night temperature on the accumulation of nonstructural carbohydrates within a cultivar (Figure 5). No significant differences in fructose, sucrose, or fructan concentrations occurred at the different temperatures. However, differences in the bulb dry-matter percentage may have been related to changes in water content influenced by these compounds. The concentration of monosac charides plus sucrose per unit of tissue water was constant in all temperature treatments, so that the osmolarity remained constant.