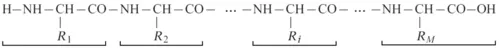Protein Physics: A Course of Lectures covers the most general problems of protein structure, folding and function. It describes key experimental facts and introduces concepts and theories, dealing with fibrous, membrane, and water-soluble globular proteins, in both their native and denatured states.
The book systematically summarizes and presents the results of several decades of worldwide fundamental research on protein physics, structure, and folding, describing many physical models that help readers make estimates and predictions of physical processes that occur in proteins.
New to this revised edition is the inclusion of novel information on amyloid aggregation, natively disordered proteins, protein folding in vivo, protein motors, misfolding, chameleon proteins, advances in protein engineering & design, and advances in the modeling of protein folding.
Further, the book provides problems with solutions, many new and updated references, and physical and mathematical appendices. In addition, new figures (including stereo drawings, with a special appendix showing how to use them) are added, making this an ideal resource for graduate and advanced undergraduate students and researchers in academia in the fields of biophysics, physics, biochemistry, biologists, biotechnology, and chemistry.
Fully revised and expanded new edition based on the latest research developments in protein physics
Written by the world's top expert in the field
Deals with fibrous, membrane, and water-soluble globular proteins, in both their native and denatured states
Summarizes, in a systematic form, the results of several decades of worldwide fundamental research on protein physics and their structure and folding
Examines experimental data on protein structure in the post-genome era
Foire aux questions
Comment puis-je résilier mon abonnement ?
Il vous suffit de vous rendre dans la section compte dans paramètres et de cliquer sur « Résilier l’abonnement ». C’est aussi simple que cela ! Une fois que vous aurez résilié votre abonnement, il restera actif pour le reste de la période pour laquelle vous avez payé. Découvrez-en plus ici.
Puis-je / comment puis-je télécharger des livres ?
Pour le moment, tous nos livres en format ePub adaptés aux mobiles peuvent être téléchargés via l’application. La plupart de nos PDF sont également disponibles en téléchargement et les autres seront téléchargeables très prochainement. Découvrez-en plus ici.
Quelle est la différence entre les formules tarifaires ?
Les deux abonnements vous donnent un accès complet à la bibliothèque et à toutes les fonctionnalités de Perlego. Les seules différences sont les tarifs ainsi que la période d’abonnement : avec l’abonnement annuel, vous économiserez environ 30 % par rapport à 12 mois d’abonnement mensuel.
Qu’est-ce que Perlego ?
Nous sommes un service d’abonnement à des ouvrages universitaires en ligne, où vous pouvez accéder à toute une bibliothèque pour un prix inférieur à celui d’un seul livre par mois. Avec plus d’un million de livres sur plus de 1 000 sujets, nous avons ce qu’il vous faut ! Découvrez-en plus ici.
Prenez-vous en charge la synthèse vocale ?
Recherchez le symbole Écouter sur votre prochain livre pour voir si vous pouvez l’écouter. L’outil Écouter lit le texte à haute voix pour vous, en surlignant le passage qui est en cours de lecture. Vous pouvez le mettre sur pause, l’accélérer ou le ralentir. Découvrez-en plus ici.
Est-ce que Protein Physics est un PDF/ePUB en ligne ?
Oui, vous pouvez accéder à Protein Physics par Alexei V. Finkelstein,Oleg Ptitsyn en format PDF et/ou ePUB ainsi qu’à d’autres livres populaires dans Biowissenschaften et Biophysik. Nous disposons de plus d’un million d’ouvrages à découvrir dans notre catalogue.
Main functions of proteins. Amino acid sequence determines the three-dimensional structure, and the structure determines the function. The reverse is not true. Fibrous, membrane, and globular proteins. Primary, secondary, tertiary, and quaternary structures of proteins. Domains. Cofactors. Active sites and protein globules. Protein biosynthesis; protein folding in vivo and in vitro. Posttranslational modifications.
Keywords
Main functions of proteins; Three-dimensional structure; Fibrous proteins; Membrane proteins; Globular proteins; Domains; Protein folding
This lecture contains an introduction to the whole course—a brief overview of what is given (or omitted) in the following lectures. Therefore, unlike other lectures, this one is not supplied with specific references; instead, it contains a list of textbooks that may be recommended for additional reading.
Proteins are molecular machines, building blocks, and arms of a living cell. Their major and almost sole function is enzymatic catalysis of chemical conversions in and around the cell. In addition, regulatory proteins control gene expression, and receptor proteins (which sit in the lipid membrane) accept intercellular signals that are often transmitted by hormones, which are proteins as well. Immunoproteins and the similar histocompatibility proteins recognize and bind “foe” molecules as well as “friend” cells, thereby helping the latter to be properly accommodated in the organism. Structural proteins form microfilaments and microtubules, as well as fibrils, hair, silk, and other protective coverings; they reinforce membranes and maintain the structure of cells and tissues. Transfer proteins transfer (and storage ones store) other molecules. Proteins responsible for proton and electron transmembrane transfer provide for the entire bioenergetics, that is, light absorption, respiration, ATP production, etc. By ATP “firing” other proteins provide for mechanochemical activities—they work in muscles or move cell elements.
The enormous variety of protein functions is based on their high specificity for the molecules with which they interact, a relationship that resembles a key and lock (or rather, a somewhat flexible key and a somewhat flexible lock). This specific relationship demands a fairly rigid spatial structure of the protein—at least when the protein is “operating” (before and after that, some proteins are “natively unfolded”). That is why the biological functions of proteins (and other macromolecules of the utmost importance for life—DNA and RNA) are closely connected with the rigidity of their three-dimensional (3D) structures. Even a little damage to these structures, let alone their destruction, is often the reason for loss of, or dramatic changes in, protein activities.
A knowledge of the 3D structure of a protein is necessary to understand how it functions. Therefore, in these lectures, the physics of protein function will be discussed after protein structure, the nature of its stability and its ability to self-organize, that is, close to the end of this course.
Proteins are polymers; they are built up by amino acids that are linked into a peptide chain; this was discovered by E. Fischer as early as the beginning of the 20th century. In the early 1950s, F. Sanger showed that the sequence of amino acid residues (a “residue” is the portion of a free amino acid that remains after polymerization) is unique for each protein. The chain consists of a chemically regular backbone (main chain) from which various side chains (R1, R2, …, Ri, …, RM) project (later on, we will consider certain deviations from the backbone,
NH
CH
CO
, regularity):
The number M of residues in protein chains ranges from a few dozens to many thousands. This number is gene-encoded.
There are 20 main (and a couple of accessory) species of amino acid residues. Their position in the protein chain is gene-encoded, too. However, subsequent protein modifications may contribute to the variety of amino acids.
Also, some proteins bind various small molecules, serving as cofactors.
In an “operating” protein, the chain is folded in a strictly specified structure. In the late 1950s, Perutz and Kendrew solved the first protein spatial structures and demonstrated their highly intricate and unique nature. However, it is noteworthy that the strict specificity of the 3D structure of protein molecules was first shown (as it became clear later) back in the 1860s, by Hoppe-Zeiler who obtained hemoglobin crystals—in a crystal each atom occupies a unique place.
The question whether the structure of a protein is the same in a crystal (where protein structures had been first established) and in a solution had been discussed for many years (when only indirect data were available) until the virtual identity of these (apart from small fluctuations) was demonstrated by nuclear magnetic resonance (NMR) spectroscopy.
Proteins “live” under various environmental conditions, which leave an obvious mark on their structures. The less water there is around, the more valuable the hydrogen bonds are (which reinforce the regular, periodic 3D structures of the protein backbone) and the more regular the stable protein structure ought to be.
According to their “environmental conditions” and general structure, proteins can be roughly divided into three classes:
1.Fibrous proteins form vast, usually water-deficient aggregates; their structure is usually highly hydrogen-bonded, highly regular, and maintained mainly by interactions between various chains.
2.Membrane proteins reside in a water-deficient membrane environment (although they partly project into water). Their intramembrane portions are highly regular (like fibrous proteins) and highly hydrogen-bonded, but restricted in size by the membrane thickness.
3.Water-soluble (residing in water) globular proteins are less regular (especially small ones). Their structure is maintained by interactions of the chain with itself (where an important role is played by interactions between hydrocarbon—“hydrophobic”—groups that are far apart in the sequence but adjacent in space) and sometimes by chain interactions with cofactors.
Finally, there are some, mostly small or hydrocarbon group-poor or charged group-rich polypeptides, which do not have an inherent fixed structure in physiological conditions by themselves but obtain it by interacting with other molecules. They are usually called “natively” (or “intrinsically”) disordered (or unfolded) proteins.
The above classification is extremely rough. Some proteins may comprise a fibrous “tail” and a globular “head” (eg, myosin), and so on.
To date, we know many millions of protein sequences (they are deposited at special computer databanks, eg, Swiss-Prot) and hundreds of thousands of protein spatial structures (they are compiled at the Protein Data Bank, or simply PDB). What we know about 3D protein structures mostly concerns water-soluble globular proteins. The solved spatial structures of membrane and fibrous proteins are relatively few. The reason is simple: water-soluble proteins are easily isolated as separate molecules, and their structure is relatively easily established by X-ray crystallography and by NMR studies in solution. That is why, when speaking about “protein structure” and “protein structure formation” one often actually means regularities shown for water-soluble globular proteins only. This must be kept in mind when reading books and papers on proteins, including these lectures. Moreover, it must be kept in mind that, for the same experimental reason, contemporary protein physics is mainly physics of small proteins, while the physics of large proteins is only starting to develop.
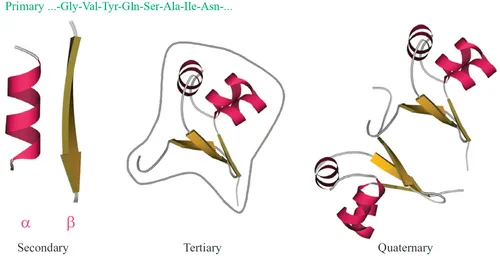
Noncovalent interactions maintaining 3D protein architecture are much weaker than chemical bonds fixing a sequence of monomers (amino acids) in the protein chain. This sequence—it is called “the primary structure of a protein” (Fig. 1.1)—results from biochemical matrix synthesis according to a gene-coded “instruction.”
Fig. 1.1 Levels of protein structure organization: primary structure (amino acid sequence); regular secondary structures (α-helix and one strand of β-structure are shown); tertiary structure of a globule formed by one chain (the gray contour outlines the body of a dense globule); and quaternary structure of an oligomeric protein formed by several chains (here, dimeric cro repressor). This figure and most of the others show schemes of the backbone folds only; an all-atom presentation of a protein is given, eg, in Fig. 1.3.
Protein architectures, especially those of water-soluble globular proteins, are complex and of great diversity, unlike the universal double helix of DNA (the single-stranded RNAs appear to have an intermediate level of complexity). Nevertheless, certain “standard” motifs are detected in proteins as well, which will be discussed in detail in the last half of this course (note that the “standard” structures are, in fact, the same ...
Table des matières
Cover image
Title page
Table of Contents
Copyright
Contents
Foreword to the First English Edition
Preface
Acknowledgements
Part I: Introduction
Part II: Elementary Interactions in and Around Proteins
Part III: Secondary Structures of Polypeptide Chains
Part IV: Protein Structures
Part V: Cooperative Transitions in Protein Molecules
Part VI: Prediction and Design of Protein Structure
Part VII: Physical Background of Protein Functions
Appendices
Problems With Solutions and Comments
Index
Normes de citation pour Protein Physics
APA 6 Citation
Finkelstein, A., & Ptitsyn, O. (2016). Protein Physics (2nd ed.). Elsevier Science. Retrieved from https://www.perlego.com/book/1832418/protein-physics-a-course-of-lectures-pdf (Original work published 2016)
Chicago Citation
Finkelstein, Alexei, and Oleg Ptitsyn. (2016) 2016. Protein Physics. 2nd ed. Elsevier Science. https://www.perlego.com/book/1832418/protein-physics-a-course-of-lectures-pdf.
Harvard Citation
Finkelstein, A. and Ptitsyn, O. (2016) Protein Physics. 2nd edn. Elsevier Science. Available at: https://www.perlego.com/book/1832418/protein-physics-a-course-of-lectures-pdf (Accessed: 15 October 2022).
MLA 7 Citation
Finkelstein, Alexei, and Oleg Ptitsyn. Protein Physics. 2nd ed. Elsevier Science, 2016. Web. 15 Oct. 2022.