![]()
PART 1
Hierarchy Theory of Evolution
![]()
General Principles of Biological Hierarchical Systems
Ilya Tëmkin and Emanuele Serrelli
The hierarchy theory of evolution is ontologically committed to the existence of inherent nested hierarchies in nature and attempts to explain natural phenomena as a product of complex dynamics of biological systems in the context of scaling. Most generally, a system is a network of functionally interdependent and structurally interconnected parts of an integrated whole, where the complexity arises from nontrivial, nonlinear interactions among parts, so that the emergent global dynamics of the whole cannot be expressed as simply the sum of its parts.
Organizing Principles of a Hierarchy
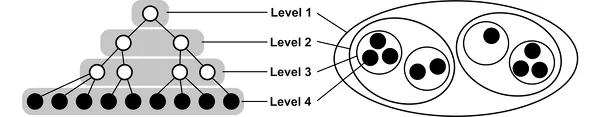
A hierarchy is an arrangement of entities according to levels, or classes of entities of the same rank or significance. The meaning of levels and the relationship among them depend on which specific type of hierarchy is considered: order, inclusion, control, or level hierarchy. Despite the fact that multiple hierarchies can be recognized in living systems (ranging from purely epistemological constructs to specific ontological claims), the hierarchy theory of evolution focuses on a particular class of hierarchies—nested compositional hierarchies—as a fundamental physical organizational principle of real biological systems. A nested compositional hierarchy is an ordered organization in the context of scale based on the principle of increasing inclusiveness, so that entities at one level are composed of parts at lower levels, which themselves function as parts of more inclusive entities at a higher level. Levels are classes of such parts, and wholes and their ranks correspond to the scale of the entities that are their members. The term focal level refers to a level at which a particular phenomenon is observed, whereas the terms higher (or upper) and lower levels refer to more inclusive and less inclusive levels relative to the focal level, respectively. A nested hierarchy can be formalized mathematically as an ordered set and represented graphically as a rooted tree (an acyclical graph) or a Venn diagram (figure P1.1).
The discreteness of levels results from differences in the nature and rate of processes between entities at a given level and entities at different levels. However, it is not always easy to draw clear boundaries between levels when there are small differences in scale among entities at levels of adjacent ranks, particularly at the more weakly integrated higher levels of a hierarchy.
The nested part-whole relation in a hierarchy produces an important distinction between two kinds of attributes or traits of entities at any given level: aggregate and emergent. Aggregate traits are cumulative properties or combined attributes of entities at the lower level (for example, stenotopic-eurytopic characteristics of a species, which derive from the cumulative range of individual tolerances to environmental changes); emergent traits are properties that cannot be reduced to or be expressed in terms of properties of entities at the lower level (such as sex ratio, which characterizes a collective state of a population that cannot be applied to individual organisms). At a given level, all entities may either share the same traits or display variation in the traits.
Hierarchical Dynamics
The interactions of entities in a hierarchy fall into two major categories: within and between levels. Within-level, high-frequency dynamics involve direct and strong, typically time-independent (reversible) interactions of entities in the same set of processes at commensurate rates. Such interactions can be effectively represented as complex networks, or systems of interacting entities that are conventionally represented by a graph, a collection of nodes (vertices) connected by links (edges). Typically, directed links describe the flow of information, matter, or energy between a source and its target(s), whereas nondirected edges show mutual interactions, where information, matter, or energy are exchanged between a pair of nodes. Network dynamics allows for undirected interactions, such as cyclical relationships and feedback loops, so that the functional interactions within such a system are temporally restricted. Consequently, entities from different hierarchical levels cannot effectively be members of the same network. In biology, networks are present at all levels of organization, from metabolism and regulation of gene expression to ecological trophic webs and social networks within populations.
Despite the differences in the rules of interactions among entities at different levels, complex biological networks—from gene expression regulatory networks to ecosystem-wide food webs—share a set of common features: they tend to be highly modular and have a high clustering coefficient, a heavy tail in the degree distribution (the frequency distribution of the number of links per node), and a short mean path length (the mean number of nodes along the shortest path between two nodes). These fundamental architectural principles result in an astonishing network-wide property that is shared by real-world biological networks across levels of biological organization. Such networks display exceptional robustness and a high degree of tolerance against random failures and external perturbations.
Substantial differences in process rates prevent entities at different levels from interacting directly, allowing for relatively weak, generally time-dependent (irreversible) interactions in an aggregate fashion. This nontransitivity of direct effects across levels is what makes the levels discrete and quasi-independent to the extent that the details of within-level interactions can be ignored when considering between-level dynamics. Such property of hierarchical dynamics allows for decoupling and investigating processes at individual levels on their own right.
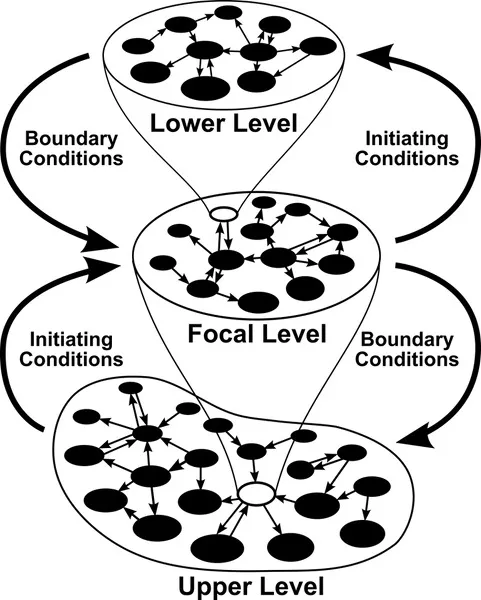
In biological systems, a pair of adjacent levels comprises a dual control system, where the interactions of entities at a lower level establish initiating conditions (upward causation) and the interactions of entities at a higher level exert constraints, or determine boundary conditions (downward causation). Noncontiguous levels may affect the dynamics at the focal level indirectly through cascading upward and downward effects across levels. However, there is a fundamental asymmetry in the effects of upward and downward causation: because downward causation affects all subsystems at all the less-inclusive levels contained within the system (level) where it originates, the dynamics at higher levels will always propagate downward, whereas the dynamics at lower levels might never manifest at higher levels.
Hierarchy and Complexity
A global architecture of a hierarchy is best approximated by a quasi-fractal representation, resulting from nesting of self-similar elements—that is, complex networks distributed across the levels—so that an entity at a focal level is composed of a network of its parts at a lower level and itself functions as an element of a network that makes up a higher level entity. Taking into account the isomorphism in the architectural features of biological networks across the hierarchy, a system of hierarchically nested, complex networks is predicted to display remarkable metastability, despite nonequilibrial dynamics at all levels of organization triggered by extrinsic disturbances, a prediction consistent with empirical data on living systems and the general tendency of complex systems to attain and remain in equilibrium. A change in such a system is a systemic, synergetic response to sufficiently strong perturbations—capable of disrupting networks at multiple levels of biological organization—that may have evolutionary consequences. Consequently, complexity in biological evolutionary phenomena arises from the synergetic effect of the network dynamics of entities at the focal level, the boundary and initiation conditions emerging from network dynamics of entities at adjacent levels, and cascading upward and downward effects from more remote levels.
The Ontological Paradigm of the Hierarchy Theory of Evolution
Living systems in general and, more specifically, evolving systems are defined by two classes of processes: interaction, or active exchange of matter and energy, and replication, or transmission of heritable information. The dynamic and informational aspects characterize two distinct kinds of biological entities: the replicators, or closed systems, that pass on their structure (i.e., information) to offspring, and the interactors, or open systems that require continuous matter/energy exchange with their environment to maintain their equilibrium. Interactors are related to replicators in such a way that the survival and the work of the former are causally responsible for the differential propagation of the latter. In evolutionary history, the tasks of replication (heritability) and dynamic interactions became progressively and irreversibly decoupled, producing differentiated and specialized sets of entities at each level of biological organization of greater functional efficiency. At the cellular level, the genetic nucleus in eukaryotes is separated from metabolic cytoplasm and other organelles; at the organismal level, the genetically transmitted germline is segregated from the mortal soma; and at the population level—for instance, in eusocial insects—nonreproductive castes are distinct from reproductive individuals. The degree of the decoupling and hence interdependency between the interactors and replicators varies substantially across organizational levels.
This fundamental dualism of biological systems and the lack of one-to-one correspondence of replicator and interactor entities across levels of biological organization prevented accommodating the entirety of the biotic realm by a single hierarchy. The hierarchy theory of evolution adopts a model of two interconnected systems, corresponding to the dynamic and the informational aspects of life: (1) the economic, or ecological, compositional nested hierarchy that captures for dynamic interactions of entities within and across levels through upward and downward causation and (2) the genealogical, or reproductive, nested compositional hierarchy, which reflects the historical nature of biological systems stemming from the unidirectional control of information flow. Most generally, the economic and genealogical hierarchies represent, respectively, the spatial and temporal dimensions of the organic realm. Importantly, drawing an explicit distinction between the two types of hierarchies allows for elucidating causal relationships between them.
References
Albert, Réka, and Albert L. Barabási. 2002. “Statistical Mechanics of Complex Networks.” Reviews of Modern Physics 74: 47–97.
Brooks, Daniel R., and E. O. Wiley. 1988. Evolution as Entropy: Toward a Unified Theory of Biology, 2nd ed. Chicago: University of Chicago Press.
Eldredge, Niles, and Stanley N. Salthe. 1984. “Hierarchy and Evolution.” Oxford Surveys in Evolutionary Biology 1: 184–208.
Hull, David. 1980. “Individuality and Selection.” Annual Review of Ecology and Systematics 11: 311–22.
Newman, M. E. J. 2003. “The Structure and Function of Complex Networks.” SIAM Review 45: 167–256.
Pattee, Howard H. 1970. “The Problem of Biological Hierarchy.” In Toward a Theoretical Biology 3, Drafts, edited by Conrad H. Waddington, 117–36. Edinburgh: Edinburgh University Press.
———, ed. 1973. Hierarchy Theory: The Challenge of Complex Systems. New York: G. Braziller.
Pumain, Denise, ed. 2006. Hierarchy in Natural and Social Sciences. Dordrecht: Springer.
Salthe, Stanley N. 1985. Evolving Hierarchical Systems: Their Structure and Representation. New York: Columbia University Press.
Simon, Herbert A. 1962. “The Architecture of Complexity.” Proceedings of the American Philosophical Society 106: 467–82.
———. 1973. “The Organization of Complex Systems.” In Hierarchy Theory: The Challenge of Complex Systems, edited by Howard H. Pattee, 1–27. New York: G. Braziller.
Strogatz, Steven H. 2001. “Exploring Complex Networks.” Nature 410: 268–76.
Tëmkin, Ilya, and Niles Eldredge. 2015. “Networks and Hierarchies: Approaching Complexity in Evolutionary Theory.” In Macroevolution: Explanation, Interpretation and Evidence, edited by Emanuele Serrelli and Nathalie Gontier, 183–226. New York: Springer.
Wimsatt, William C. 1976. “Reductionism, Levels of Organization, and the Mind-Body Problem.” In Consciousness and the Brain: A Scientific and Philosophical Inquiry, edited by Globus Gordon, G. Maxwell, and I. Savodnik, 205–67. New York: Plenum Press.
Woodger, J. H. 1929. Biological Principles: A Critical Study. London: Kegan Paul, Trench, Trubner.
![]()
Summaries for Part 1
The contributions in this part address conceptual and terminological questions that received little attention in evolutionary theory and philosophy of biology but are of critical importance for the unification of evolutionary biology in general and the hierarchy theory of evolution in particular.
Bruce Lieberman tackles an epistemological issue about evolutionary biology in hierarchical perspective: Is it a science discovering mechanisms (a nomothetic science) or is it a historical science (an idiographic one)? Lieberman reconstructs the story of this distinction in paleobiology, showing that an opposition between the two kinds is not necessary. Epistemological proposals to unify nomothetic and idiographic sciences have been convincing. In fact, macroevolutionary theories such as punctuated equilibria, the turnover pulse hypothesis, and the sloshing bucket theory embody both aspects. They invoke unique historical events, contingent factors, and deterministic mechanisms to generate and explain repeated patterns. Lieberman argues that “at least from the perspective of macroevolutionary theory, repeated analysis of contingent histories is the key to discovering whatever nomothetic principles exist in the history of life.”
Gustavo Caponi addresses a fundamental distinction between two kinds of biological individuals: systems and lineages. This chapter attempts to overcome the disconnect in logic that is traditionally applied in the analysis of these two kinds of individuals, leveling the ground for a unified theoretical perspective. To this aim, the chapter focuses on the “modes of incorporation” that differentiate systems and lineages and on causal integration, emphasizing that systemic causality does not apply to lineages, where causal integration is due to genealogical relationships. Further discussion focuses on the levels of individual organism and population (both simultaneously characterized by properties of systems and lineages) to examine the interaction between the ecological and the genealogical aspects of biological hierarchies. In light of the system/lineage dichotomy, an important distinction is made between the notions of character and part.
Jon Umerez proposes an articulated view of hierarchical levels that distinguishes five kinds of progressively restrictive relationships among elements, leading to the emergence of a level: composition, integration, emergence, r...