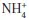Amino acids play a role in the defence mechanisms and stress responses of plants, as well as in food quality and safety for humans and animals. Recent advances in the field make a comprehensive overview of the information a necessity; this book collates chapters on plant enzymes and metabolism, modulation, molecular aspects and secondary products. Also including information on ecology, the environment and mammalian nutrition and toxicology, it provides an authoritative resource.
Foire aux questions
Comment puis-je résilier mon abonnement ?
Il vous suffit de vous rendre dans la section compte dans paramètres et de cliquer sur « Résilier l’abonnement ». C’est aussi simple que cela ! Une fois que vous aurez résilié votre abonnement, il restera actif pour le reste de la période pour laquelle vous avez payé. Découvrez-en plus ici.
Puis-je / comment puis-je télécharger des livres ?
Pour le moment, tous nos livres en format ePub adaptés aux mobiles peuvent être téléchargés via l’application. La plupart de nos PDF sont également disponibles en téléchargement et les autres seront téléchargeables très prochainement. Découvrez-en plus ici.
Quelle est la différence entre les formules tarifaires ?
Les deux abonnements vous donnent un accès complet à la bibliothèque et à toutes les fonctionnalités de Perlego. Les seules différences sont les tarifs ainsi que la période d’abonnement : avec l’abonnement annuel, vous économiserez environ 30 % par rapport à 12 mois d’abonnement mensuel.
Qu’est-ce que Perlego ?
Nous sommes un service d’abonnement à des ouvrages universitaires en ligne, où vous pouvez accéder à toute une bibliothèque pour un prix inférieur à celui d’un seul livre par mois. Avec plus d’un million de livres sur plus de 1 000 sujets, nous avons ce qu’il vous faut ! Découvrez-en plus ici.
Prenez-vous en charge la synthèse vocale ?
Recherchez le symbole Écouter sur votre prochain livre pour voir si vous pouvez l’écouter. L’outil Écouter lit le texte à haute voix pour vous, en surlignant le passage qui est en cours de lecture. Vous pouvez le mettre sur pause, l’accélérer ou le ralentir. Découvrez-en plus ici.
Est-ce que Amino Acids in Higher Plants est un PDF/ePUB en ligne ?
Oui, vous pouvez accéder à Amino Acids in Higher Plants par J P F D'Mello, J D'Mello en format PDF et/ou ePUB ainsi qu’à d’autres livres populaires dans Biological Sciences et Botany. Nous disposons de plus d’un million d’ouvrages à découvrir dans notre catalogue.
Glutamate dehydrogenase (GDH; EC.1.4.1.2) aminating activity unleashes a metabolic propulsion in crop plants that optimizes, maximizes and doubles the amino acid, protein, fatty acid, carbohydrate and dry matter yields per hectare. It is possible that the molecular biology of the enzyme may hold clues for increasing the yields of food crops for feeding a burgeoning world population without cultivating more land, without applying more fertilizer and without increasing man-hour input. GDH is historically known to catalyse the reversible reductive amination of α-ketoglutarate (α-KG) in the synthesis of L-glutamate in higher plants. It also catalyses the template-independent polymerization of nucleoside triphosphate(s) to RNA. The discovery of the aminating activity of the glutamine synthetase (GS; EC6.3.1.2)–glutamate synthase (GOGAT; EC.2.6.1.53) cycle in chloroplasts triggered new conversations and extensive research on the role of GDH in the synthesis of glutamate. The Km value for the
ion is much lower in GS than in GDH, which fact has increased understanding of the differential mechanisms of
ion assimilation and glutamate synthesis during deficit and surplus supplies of α-KG. It might appear from some repetitive experimentation on low-activity preparations of GDH that the deaminating direction is emerging as the consensus role of the enzyme. But many lines of new experimental evidence from systems biology illuminate the uninterruptible flow of α-KG via the citric acid cycle; and the nature of the aminating cassette of GDH isoenzymes reiterates and supports the amination role. Specifically, the kinetic property of GDH to discriminate and integrate the plant’s environmental signals by synthesizing RNAs that regulate the mRNAs encoding many enzymes (phosphate translocator, granule-bound starch synthase, phosphoglucomutase, nitrate reductase, glycinamide ribonucleotide transformylase, etc.) of primary metabolism is the collateral chemical scaffolding on which the aminating natural role stands as it synchronizes carbon and nitrogen metabolic pathways, and assimilates exponential multiples of kilogram quantities of
ion as assimilated via the GS-GOGAT cycle during normal to maximum photosynthetic supplies of C3, C5, etc. compounds. GDH possibly deaminates vanishing fractions of glutamate as it synthesizes RNA that regulates the mRNAs so that glutamate biosynthesis is not shut down during low to normal flow of C3, C5, etc. compounds of photosynthesis. Ammonium ion assimilation and glutamate synthesis by GDH are regulated in higher plants by the supply of
ions, and the flow of α-KG from the citric acid cycle to GDH, not by the reverse-flow of α-KG from glutamate.
1.2 Introduction
L-Glutamic acid occupies the central position in amino acid metabolism because other amino acids (except lysine) can be derived from it when it transfers its amino group to their α-keto acids (D’Mello, 2003). Glutamate dehydrogenase (L-glutamic acid:GDH NADH oxidoreductase; EC.1.4.1.2) is the pre-eminent enzyme (Strecker, 1953; Pahlich et al., 1980; Magalhaes, 1991) known to catalyse the reversible reductive amination of α-ketoglutarate (α-KG) in the synthesis of glutamate in all higher plants studied. It also catalyses template-independent polymerization of nucleoside triphosphate(s) to RNA (Osuji et al., 2003a). The discovery of the aminating activity of glutamine synthetase (GS; EC6.3.1.2)-glutamate synthase (GOGAT; EC.2.6.1.53) cycle in chloroplasts (Lea and Miflin, 1974) triggered new conversations and intensive research on the role and regulation of GDH in the assimilation of
ions and synthesis of glutamate. The Km value for the
ion is much lower in GS than in GDH, which fact has increased the understanding of the differential mechanisms of
ion assimilation and glutamate biosynthesis during normal and surplus supplies of α-KG (Robinson et al., 1992; Osuji and Madu, 2012). This review discusses recent experimental evidence on the interwoven natural roles of GDH in glutamate synthesis, the coordination of nitrogen and carbon metabolic pathways, and glutamate deamination. GDH is multi-isoenzymatic and isomerizes in response to a wide assortment of genetic, biotic and abiotic environmental factors. Several experiments have focused narrowly and repeatedly on a handful of low activity GDH preparations while neglecting the natural aminating role of the preponderantly high activity GDH preparations. It might appear from such repetitive narratives on the handful of the same experimental approach that the consensus role of GDH is moving towards the deamination of glutamate (Robinson et al., 1991; Glevarec et al., 2004; Lehmann and Ratajczak, 2008; Labboun et al., 2009). The approach of this review is experimentally different, being focused on the molecular biology of GDH amination and deamination reactions (Yamaya et al., 1984; Oaks, 1994; Melo-Oliveira et al., 1996; Pahlich, 1996; Syntichaki et al., 1996; Madu, 1997; Turano et al., 1997; Osuji and Braithwaite, 1999; Ameziane et al., 2000; Pavesi et al., 2000), especially those that explain the biochemistry of the isomerization reaction in crops treated with pesticides, mineral fertilizer, N-carboxymethylchitosan and plant hormones; and synchronizing the primary nitrogen and carbon metabolic pathways of citric acid cycle, glycolysis, photosynthesis and nitrogen assimilation, leading to maximum yields of glutamate and other amino acids, proteins, carbohydrates, fatty acids, etc. (Osuji and Madu, 2012). Some of the incomplete arguments embedded in earlier conversations on the roles of the enzyme will be highlighted. The review discusses many lines of new experimental evidence from systems biology (metabolism) that illuminate the uninterruptible flow of C3, C4, C5, etc. compounds; the nature of the aminating cassette of GDH isoenzymes; the large kilogram quantities of amino acids and proteins that result from GDH amination activity; and the application of the GDH amination role to increase the yields of maize, roots/tubers and leguminous crops for combating kwashiorkor (protein energy malnutrition) and food insecurity in some arid regions.
1.3 Glutamate Dehydrogenase Structure and Localization
GDH has a hexameric structure of three subunit polypeptides controlled by two non-allelic genes GDH1 and GDH2, responsible for the synthesis of subunits ‘a’ and ‘b’ respectively; with gene GDH1 synthesizing subunits ‘a’ and ‘α’, two codominant allelic forms; and GDH2 synthesizing the ‘b’ subunit (Cammaerts and Jacobs, 1983). The binomial combination of the three subunits into hexamers gives the ~28 isoenzyme distribution typically obtained in 2-D electrophoretic resolution of the enzyme extracted from soybean (Osuji et al., 2003b), peanut (Osuji and Braithwaite, 1999), sweet potato (Osuji and Madu, 1997), maize (Osuji and Madu, 1995), yam tuber (Osuji et al., 1991) and many other higher plants. Since isomerization is the committed step in the oxidoreductase catalytic mechanism of GDH (Osuji and Braithwaite, 1999), the activities of the enzyme changing with the alterations in the isoenzyme pattern (Cammaerts and Jacobs, 1985), it is necessary in all experimentation with the enzyme to demonstrate the qualitative alterations in the isoenzyme pattern (Osuji and Madu, 1996; Watanabe et al., 2011). Presentation of the GDH isomerization profiles inspires confidence in the quality assurance evaluation, in the baseline for validating the experimental biological tissues, and authenticates the quality control measures practised during the instrumental analytical data generation. In this regard, the free solution (Rotofor) isoelectric focusing (IEF) of GDH, followed by native PAGE of the fractions, introduced a considerable advancement in the analytical chemistry of the enzyme (Osuji and Braithwaite, 1999; Osuji et al., 1999) because the isoenzymes become fully separated one from the other, being elegantly projected in three dimensions in some cases. It is noteworthy, therefore, that all the conversations that withheld the aminating GDH isoenzyme profiles have invariably supported the deaminating function of the enzyme (Miflin and Lea, 1976; Robinson et al., 1992, 1992; Stewart et al., 1995; Sadunishvili et al., 1996; Aubert et al., 2001; Miflin and Habash, 2002; Glevarec et al., 2004; Masclaux-Daubresse et al., 2006; Purnell and Botella, 2007; Skopelitis et al., 2007; Miyashita and Good, 2008; Labboun et al., 2009).
Nucleotide sequences of GDH cDNA and deduced amino acid sequences of GDH polypeptides (Syntichaki et al., 1996; Pavesi et al., 2000) have been studied as well as the protein structure of hexameric GDH (Britton et al., 1992). The highest level of sequence similarities in GDHs were found within the N-terminal half of the polypeptides, the region thought to be involved in oxoglutarate/glutamate binding and catalytic activity. Analyses of the conserved residues in 3-D projection attest to a remarkable similarity in the layout of the active site of GDHs. Specifically, in the hexamer, each subunit polypeptide is folded into two domains separated by a deep cleft which harbours the NADH binding site. Adjacent to the domain of the NADH is a strategic cluster of lysine residues, one of which forms the Schiff base intermediate and facilitates the transfer of the hydrogen of the nicotinamide (Brown et al., 1973) to α-iminoglutarate in the catalytic reduction reaction (Osuji et al., 1999). These structural details support the non-redundant role of the enzyme in the reversible amination of α-KG (Melo-Oliveira et al., 1996).
GDH is easily found in mitochondria (Yamaya et al., 1984; Calle et al., 1986; Bravo and Maeso, 1987). GDHs have also been identified in chloroplasts (Turano et al., 1996) and, earlier, in the cytosol (Chou and Splittstoesser, 1972). These strategic localizations, especially in the cytosol and mitochondria...
Table des matières
Cover
Half Title
Title
Copyright
Contents
Contributors
Preface
Glossary
PART I ENZYMES AND METABOLISM
PART II DYNAMICS
PART III CHEMICAL ECOLOGY
PART IV PLANT PRODUCTS: QUALITY AND SAFETY
PART V CONCLUSIONS
Index
Normes de citation pour Amino Acids in Higher Plants
APA 6 Citation
[author missing]. (2015). Amino Acids in Higher Plants ([edition unavailable]). CABI. Retrieved from https://www.perlego.com/book/969532/amino-acids-in-higher-plants-pdf (Original work published 2015)