![]()
Capitolo 1
Equivoci intorno alla genetica: gemelli, jazz e topi agouti
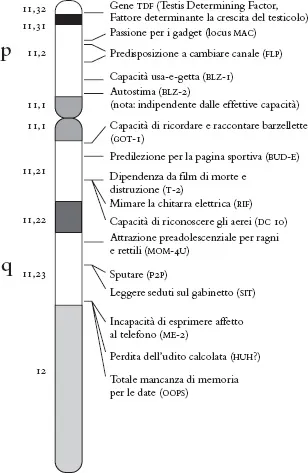
Come la genetica possa dare origine a distorsioni nell’interpretazione del suo reale ruolo nel condizionare i comportamenti umani è suggerito dalla figura di p.4, che attribuisce al cromosoma Y (quello maschile) un ruolo causale per una serie di comportamenti più o meno aberranti considerati tipici del maschio medio: la capacità di ricordare e raccontare barzellette, sputare, leggere seduti sul gabinetto, l’incapacità di esprimere affetto al telefono… Naturalmente si tratta di uno scherzo, che da un lato riflette un evidente abuso, nei mezzi di informazione di massa, della capacità della genetica di spiegare i comportamenti e le malattie, e dall’altro… beh, dall’altro sembra cogliere qualche caratteristica distintiva dell’universo maschile, non sappiamo quanto attribuibile al cromosoma Y e quanto all’ambiente o all’educazione.
Gemelli e figli adottivi
Le polemiche sulla genetica e la manipolazione genetica sono viziate da un malinteso di fondo, peraltro alimentato dagli stessi scienziati quando hanno insistito sulla centralità del DNA, il Sacro Graal della biologia moderna. Il DNA è sicuramente centrale, ma molto meno di quanto si pensasse dieci o venti anni fa. I geni contengono le istruzioni per lo sviluppo e per il corretto (o scorretto) funzionamento dell’organismo, ma gran parte di ciò che realmente si realizza del progetto originario – cioè come il genotipo diventa fenotipo – dipende largamente da elementi esterni al DNA e alla stessa cellula, in altre parole dipende dall’ambiente. Vediamo alcuni dei motivi che oggi fanno ridimensionare l’importanza del DNA.
La ricerca sui gemelli è stata avviata da più di un secolo (i primi studi scientifici importanti risalgono almeno a Francis Galton, cugino di Darwin, nel 1875), in base all’assunto che i gemelli, in particolare quelli monozigoti, cioè geneticamente identici, si ammalino delle stesse malattie avendo lo stesso patrimonio ereditario. Un altro filone importante è costituito dal confronto tra figli biologici e figli adottivi: se è la genetica a condizionare l’insorgenza delle malattie, ci aspettiamo che i figli biologici tendano a condividere più spesso le stesse cause di morte dei genitori rispetto a quelli adottivi (che hanno un diverso patrimonio genetico). Sorensen e colleghi hanno trovato, contrariamente alle attese, che i figli biologici, paragonati a quelli adottivi, tendono a condividere in modo molto blando le cause di morte dei loro genitori, con l’eccezione delle malattie infettive (Sorensen, 1998). Questo risultato è piuttosto sorprendente, perché a priori ci si attendeva che vi fosse una maggiore predisposizione familiare per le malattie cardiovascolari, neurologiche o neoplastiche. Anche i risultati delle ricerche sui gemelli sono stati meno ovvi dell’atteso. In un grande studio prospettico, sono state identificate coppie di gemelli monozigoti in cui entrambi erano non-fumatori, altre in cui entrambi fumavano, e altre in cui uno solo fumava. Ebbene, i tumori al polmone che si manifestarono erano inequivocabilmente attribuibili al fumo, poiché l’incidenza di cancro era molto più alta nei fumatori che nei non-fumatori, indipendentemente dalla gemellarità (Jongbloet, 1997).
Fig.1. La localizzazione di presunti geni del comportamento maschile nel cromosoma Y. Da http://www.bioteach.ubc.ca/TeachingResources/Genetics/Ychromosome.pdf
Tra i gemelli monozigoti (identici), circa due terzi possiedono una placenta in comune, perché la divisione in due organismi separati è avvenuta dopo la formazione del chorion, la parte embrionale della placenta. Il restante terzo di gemelli identici è costituito da gemelli che possiedono due placente distinte. Più di 20 anni fa Melnick e colleghi mostrarono che i gemelli identici che avevano la placenta in comune avevano una probabilità più alta di avere lo stesso quoziente di intelligenza di quanto non avvenisse per gemelli con diverse placente, pur geneticamente identici. Questi risultati, confermati da studi più recenti, suggeriscono chiaramente che l’ambiente prenatale gioca un ruolo importante, al di là delle caratteristiche genetiche (Moore, 2001). Molte indagini hanno confrontato gemelli identici (monozigoti) con gemelli dizigoti (geneticamente diversi), osservando che i primi erano tra loro più simili dei secondi e giungendo erroneamente alla conclusione che pertanto i geni erano alla base delle differenze; tuttavia, tali studi non consideravano che mentre tutti i gemelli dizigoti hanno diverse placente, circa due terzi dei gemelli monozigoti condividono la placenta, e pertanto hanno anche lo stesso ambiente intrauterino in comune. Ma c’è di più: indagini sperimentali su uno dei più semplici organismi pluricellulari, un nematode dotato di sole 1031 cellule, hanno mostrato che individui identici dal punto di vista genetico e vissuti esattamente nello stesso ambiente artificiale sviluppano marcate differenze comportamentali; in altre parole, il caso sembra giocare una parte non indifferente, pur in condizioni sperimentali molto controllate (Moore, 2001).
Un altro ovvio espediente per studiare il ruolo della genetica nei confronti dell’ambiente consiste nello studiare le popolazioni che emigrano: se la causa delle malattie è la genetica, il rischio di contrarre malattie dovrebbe rimanere stazionario nei migranti rispetto a chi è rimasto in patria. Invece non è così. L’effetto della migrazione è addirittura drammatico per certi tipi di tumori, come i tumori del colon e i melanomi, manifestandosi già nella prima generazione di migranti. Per esempio, negli inglesi emigrati in Australia è stata osservata una chiara relazione (un gradiente) del rischio con la durata del soggiorno in Australia, e lo stesso è stato osservato per i tumori del colon nei giapponesi emigrati negli Stati Uniti. In questi casi l’emigrazione avveniva da aree a basso rischio ad aree ad alto rischio. Ma è stato osservato anche l’inverso. I migranti da aree ad alto rischio mostrano una riduzione del rischio di cancro dello stomaco e del fegato quando emigrano in aree a basso rischio, e questo fenomeno sembra dipendere strettamente dall’età all’emigrazione. Nel caso del cancro dello stomaco e del fegato, l’esposizione rispettivamente all’Helicobacter pylori (un batterio) o al virus dell’epatite B o C nell’infanzia stabilisce il rischio di base, tipico delle aree di origine, mentre la ridotta esposizione a cofattori spiega l’effetto della migrazione in aree a basso rischio. Uno dei cofattori più importanti per i tumori del fegato è l’aflatossina, un contaminante di diversi cibi nei paesi in via di sviluppo.
In sostanza, le ricerche sui figli adottivi, sui gemelli e sui migranti suggeriscono chiaramente che l’ambiente conta molto più dei geni nel causare i tumori e le altre malattie croniche oggi dominanti.
I limiti del determinismo genetico – della tendenza cioè ad attribuire ai geni la maggiore responsabilità nelle variazioni di caratteri come l’intelligenza o la predisposizione alle malattie – sono stati messi in luce molto chiaramente qualche anno fa nel dibattito intorno all’infausto volume The Bell Curve di Herrnstein e Murray, che sosteneva che le differenze nel quoziente di intelligenza (QI) tra bianchi e neri americani avevano cause genetiche. La critica più lucida a questo libro è forse quella di Block (1995), basata su un precedente articolo di Richard Lewontin. Block fa notare che «ereditabilità» e «meccanismo genetico» sono due fenomeni ben distinti. La prima esprime quanto, della variazione di un carattere nella popolazione, può essere attribuito a caratteristiche già presenti nei genitori, mentre il secondo fa riferimento alla struttura del DNA, dei geni. Due esempi servono meglio di tanta teoria: (a) negli anni Cinquanta si sarebbe potuto dire che portare orecchini veniva trasmesso ereditariamente dalle madri alle figlie femmine (nessun maschio portava allora orecchini), cioè il tratto aveva una ereditabilità quasi del 100% nelle donne!; anche portare la gonna, ancora oggi, ha una forte ereditabilità di tipo matrilineare, con l’eccezione degli scozzesi; (b) il numero di dita nella specie umana è 5 e questo numero viene indubbiamente regolato dai geni (dal DNA), ma l’ereditabilità di 4 o 6 dita è molto bassa: i rari esempi di dita in più o in meno sono dovuti a eventi che occorrono durante la gravidanza, come l’esposizione al farmaco teratogeno talidomide. Pertanto gli orecchini sono ereditabili ma non attraverso i geni, e il numero di dita è determinato dai geni ma la sua variazione non è ereditabile. Come si spiega questo paradosso? Semplicemente con il fatto che noi ereditiamo tante cose dai nostri genitori, non solo i geni, ma anche l’ambiente in cui nasciamo e cresciamo, i comportamenti, e così via; e d’altra parte i geni regolano la fisiologia in un ambiente normale, ma mutamenti ambientali – come l’esposizione a talidomide – modificano l’esito della regolazione genica. Questa semplice distinzione tra orecchini e numero di dita dovrebbe aiutare a dissolvere tante false discussioni sul “gene” della depressione, dell’omosessualità, ecc. C’è anche un altro argomento più sottile e sofisticato intorno alla polemica natura-cultura, e cioè il fatto che un certo insieme di geni ereditato dai genitori viene in realtà ereditato in un particolare ambiente, per il quale è stato selezionato: in altre parole, la prole eredita certo un insieme di geni, ma anche l’ambiente per il quale quei geni sono stati selezionati, o con il quale essi si sono coevoluti. Pertanto separare le due componenti nell’interazione geni-ambiente è concettualmente spesso molto difficile.
Le obiezioni alla tesi specifica di Herrnstein e Murray circa i rapporti tra quoziente di intelligenza e razza sono decisive. Primo, il modo stesso in cui è misurato il QI è molto più l’espressione di differenze culturali ed educative che di fenomeni biologici. Secondo, il QI è cresciuto di 21 punti in Olanda tra il 1952 e il 1982, una differenza superiore a quella riscontrata negli Stati Uniti tra bianchi e afroamericani. Terzo, in diverse parti del mondo vi sono minoranze che hanno scarso accesso alla scuola e agli strumenti culturali, e hanno un QI inferiore; ma quando individui appartenenti a queste minoranze emigrano, regolarmente acquisiscono un QI superiore e più prossimo a quello delle popolazioni presso cui emigrano. Si tratta di un fenomeno del tutto analogo all’effetto della migrazione sul rischio di tumori e di malattie cardiovascolari, e che costituisce uno dei più potenti argomenti a favore di un’origine ambientale e non genetica di queste condizioni.
Waddington (1942) qualche decennio fa coniò un termine essenziale per capire le interazioni tra geni e ambiente, «canalizzazione». Si immagini una palla che rotola da una montagna: essa incontrerà diversi crinali che separano altrettante vallate; anche se il fatto di imboccare l’uno o l’altro versante del crinale può essere effetto del caso o delle circostanze fortuite, una volta imboccato un certo versante la palla sarà canalizzata verso quella specifica vallata. I tratti fenotipici, cioè le modalità con cui gli individui si sviluppano acquisendo le loro caratteristiche individuali, possono essere altamente canalizzati, e pertanto sembrare indipendenti dal contesto ambientale, come se si trattasse di vallate profonde che impediscono l’azione di altre forze sulla traiettoria della palla. Tuttavia, non è affatto scontato che tratti altamente canalizzati, cioè scarsamente suscettibili all’azione di cause ambientali, lo siano per cause genetiche. Un esperimento molto interessante è stato condotto da Gottlieb su una varietà di anatre. Se gli embrioni vengono esposti, durante la vita intrauterina, al richiamo vocale materno, essi risponderanno a tale richiamo (riconoscendo la loro madre) anche dopo la nascita. Ma se non sono esposti al richiamo materno, dopo la nascita essi saranno attratti da qualunque richiamo di altre specie di uccelli. Secondo gli studi di Gottlieb, l’avere udito il richiamo materno durante la vita intrauterina ha costituito una forma di canalizzazione che impedisce ai neonati di divenire sensibili ai richiami di altri uccelli. Con varie manipolazioni, egli fu in grado di indurre le giovani anatre addirittura a preferire il richiamo di altri uccelli. Tuttavia, quando gli embrioni erano esposti sia al richiamo della madre sia a quello di una gallina, preferivano dopo la nascita quello della madre, indicando che oltre alla canalizzazione vi è un fenomeno definito di buffering, di protezione della modalità più naturale (evolutivamente più favorevole) da interferenze esterne. Il buffering ha probabilmente un’origine genetica, molto più della canalizzazione.
Più in generale, l’esistenza di fenomeni come la canalizzazione e il buffering dipende sia da influenze genetiche sia da influenze ambientali e la separazione dei due tipi di influenza non è immediata. Per esempio, può essere difficile riconoscere i fattori ambientali che contribuiscono allo sviluppo di un tratto quando essi sono ubiquitari: se tutti fumassero 20 sigarette al giorno, il cancro sembrerebbe una malattia genetica, mentre invece il 90% dei tumori polmonari sono dovuti al fumo. Tratti che non sono per nulla canalizzati, per esempio lo stile della pettinatura, sono ovviamente influenzati da stimoli ambientali, ma per altri tratti l’origine può sembrare esclusivamente genetica perché le cause ambientali sono sconosciute oppure sono estremamente omogenee.
Ma ci sono altre argomentazioni, più di fondo, sul fatto che il DNA è solo uno degli attori nei processi dello sviluppo dell’organismo e della comparsa della malattie. È forse il protagonista, ma certamente non si tratta di un monologo.
Jazz, DNA e topi agouti
Uno dei malintesi riguardanti il DNA (e anche l’importanza del suo sequenziamento) è che il DNA contenga tutta l’informazione per lo sviluppo degli organismi o addirittura per il loro stato di salute e per il rischio di malattie. La metafora del DNA come programma di computer in cui sono contenute tutte le premesse per il corretto (o scorretto) funzionamento del corpo, e magari della psiche, è alla base di un gran numero di malintesi e di aspettative mal riposte. Piuttosto, una metafora più realistica che è stata proposta è quella del «copione» teatrale (Lewis, 1999), per cui l’espressione fenotipica può essere vista come un’interpretazione, una «performance». Ricordo che il genotipo è ciò che è inscritto nei geni, il messaggio, mentre il fenotipo è ciò che appare all’esterno, la concreta manifestazione del genotipo. Il fenotipo sarebbe dunque «la rappresentazione del copione, che porta il testo a una vita unica e vibrante proprio come gli attori sul palcoscenico». In realtà, una metafora forse più potente, per diversi motivi che chiarirò, è quella del jazz.
La sequenza di nucleotidi, cioè dei mattoni di cui è costituito un gene, può essere vista come uno spartito. Spesso il jazz ha usato e usa vecchie ballate popolari o canzoni dei musical di Broadway, per esempio My Favourite Things. Nella grande maggioranza dei casi, le interpretazioni delle ballads si basano sulla trasmissione del tema da un musicista all’altro nel corso di sessioni. Inevitabilmente – e questo peraltro fa parte del linguaggio del jazz – il tema originario viene stravolto fino a diventare quasi irriconoscibile. (L’improvvisazione nel jazz, in realtà, segue regole molto più strette di quanto il profano non pensi; la differenza con le regole come vengono abitualmente intese è che le regole dell’improvvisazione non possono essere chiaramente definite. Il fatto che Charlie Parker o Miles Davis seguano regole implicite è evidente dalla unicità e al contempo coerenza dei loro rispettivi stili e delle loro interpretazioni; nonostante ogni sforzo, tradurre in concetti le regole implicite delle loro interpretazioni, che pure esistono, si è rivelato molto difficile.)
Lo stile di uno specifico musicista include almeno quattro componenti: il tema originario, spesso un semplice pretesto; la sua personale storia, la musica che ha ascoltato e i musicisti che l’hanno influenzato; un particolare talento individuale, uno stile inimitabile; e infine l’estro del momento, basato su eventi casuali, l’umore momentaneo, l’ambiente.
Che cosa ha a che fare tutto questo con il DNA? Le similitudini sono numerose. In entrambi i casi c’è uno script, un copione, ma quel che conta, il fenotipo, è largamente determinato dall’ambiente. Il messaggio contenuto in una sequenza di DNA è soggetto a trascrizione (un termine che ricorre, notoriamente, in campo musicale), cioè viene tradotto in RNA che a sua volta viene trasformato in una proteina. Mentre fino a pochi anni fa uno dei dogmi della biologia era un gene-una proteina, si riteneva cioè che vi fosse una corrispondenza semplice e univoca tra una sequenza di DNA (un gene) e la corrispondente proteina, la scoperta dell’alternative splicing ha aperto un mondo nuovo. Ogni sequenza di DNA, infatti, può essere trascritta in diversi modi alternativi: un pezzo può essere usato insieme a una sequenza adiacente per produrre una certa proteina, un altro pezzo per produrre una seconda proteina, e così via. La trascrizione, insomma, si basa su una interpretazione del messaggio (dello spartito) che varia a seconda del momento e delle necessità. Questo consente di disporre di un numero molto maggiore di risorse (di proteine codificabili) a partire da un numero di geni relativamente ridotto (25 000). Dunque il primo punto è l’interpretazione dipendente dal contesto.
Un secondo punto è emerso solo recentemente, ed è costituito dai cambiamenti definiti epigenetici, cioè non consistenti in una mutazione, in un cambiamento strutturale del DNA, ma in un mutamento funzionale trasmissibile alle generazioni future. Nonostante l’interpretazione darwiniana – e non quella lamarckiana – sia emersa come vincente sul lungo periodo, ovvero i caratteri acquisiti non si trasmettono alle generazioni successive in un modo meccanico e banale (la giraffa non ha il collo lungo a forza di sporgersi verso le foglie più alte, trasmettendo il carattere alle generazioni successive), in un senso più debole e più sofisticato una certa forma di lamarckismo si sta rivelando complementare al darwinismo. In sostanza, sta emergendo come estremamente potente nel modulare gli effetti dell’ambiente sul ruolo del DNA il meccanismo della metilazione: gli stimoli ambientali sono infatti in grado di indurre una metilazione di tratti del DNA che comportano la trascrizione (espressione) o meno dei geni corrispondenti (la metilazione è la cessione di gruppi chimici metilici). Durante lo sviluppo dell’embrione il pattern d...