Le cerveau fait partie du corps. Il en partage la destinée, puisque c’est par le corps qu’il entre en contact avec le monde extérieur, acquiert son expérience et se construit pendant le développement et tout au long de la vie. C’est aussi par le corps que le cerveau agit sur le monde extérieur, en guidant les actions, en influençant le cours des événements. Enfin, le cerveau est en interaction permanente avec l’ensemble des organes qui composent le corps. Il régule leur fonctionnement et subit à son tour leur influence. Ce chapitre explore les relations entre le cerveau et le corps d’un double point de vue : celui du corps interface avec le monde extérieur, et celui du corps compagnon dans un même monde intérieur.
Le cerveau, le corps et le monde extérieur
Le corps est immergé dans un champ de forces physiques. Il possède des capteurs, les organes des sens, qui lui permettent d’enregistrer les variations de ces forces : les ondes lumineuses sont captées par la rétine, les ondes sonores par la cochlée, les forces mécaniques (de contact, de frottement ou de pression) et les radiations thermiques par des récepteurs situés dans la peau. Enfin, des capteurs sensibles à la forme de certaines molécules sont à la base des sens chimiques comme l’olfaction et le goût. Le type et la forme des capteurs présents dans nos organes des sens limitent la gamme des variations du monde physique que nous pouvons enregistrer. De ce fait, certains phénomènes physiques nous échappent totalement. D’autres espèces, possédant des capteurs différents, sont capables d’enregistrer ces phénomènes : l’oreille de la chauve-souris entend les ultrasons, les capteurs thermiques de certains reptiles enregistrent les radiations en infrarouge et les oiseaux migrateurs sont sensibles aux variations du champ magnétique terrestre.
Les organes des sens ont pour fonction de traduire les variations physiques et chimiques provenant du monde extérieur recueillies par ces capteurs en impulsions nerveuses, ce qu’on appelle la « transduction sensorielle ». Dans la rétine, la transduction est une opération photochimique : des pigments photosensibles situés à l’intérieur des cellules réceptrices (cônes et bâtonnets) subissent une transformation de leur structure moléculaire suite à l’absorption de la lumière. Cette transformation agit sur les canaux ioniques situés dans la membrane de la cellule, ce qui dépolarise cette dernière et provoque finalement la libération du neurotransmetteur. Dans l’appareil olfactif, les molécules odorantes de l’air s’accouplent avec des récepteurs situés dans la muqueuse du nez. La douleur résulte de l’excitation chimique de petites arborescences nerveuses présentes dans pratiquement tous les tissus de l’organisme par des molécules étrangères résultant de la lésion des tissus. Ailleurs, il s’agit de déformations mécaniques des capteurs : pour l’audition, par exemple, l’alternance d’ondes de compression et de dépression de l’air produite par les vibrations sonores provoque la vibration du tympan et des osselets de l’oreille moyenne. Ces vibrations entraînent des variations de pression du liquide qui baigne l’oreille interne ce qui, en définitive, déforme les cellules ciliées de la rampe tympanique. La déformation des cellules modifie leur potentiel de membrane. Quant aux sensations tactiles, elles sont dues à la déformation directe des capteurs provoquées par le contact des objets avec la peau.
Les informations nerveuses produites par plusieurs organes des sens à propos d’un même objet fusionnent pour en donner une représentation globale. Prenons comme exemple l’analyse sensorielle d’un aliment placé dans la bouche. Ses propriétés chimiques sont analysées simultanément par les récepteurs du goût et ceux de l’odorat : sous l’influence de la mastication, des corps volatils s’en échappent et gagnent les récepteurs du nez par l’arrière de la bouche. La combinaison du goût et de l’odorat aboutit à déterminer l’arôme de l’aliment. En même temps, les récepteurs tactiles situés à l’intérieur de la bouche donnent des informations sur sa consistance (onctuosité, velouté, etc.) et sa température ; des récepteurs mécaniques situés dans les muscles masticateurs renseignent sur sa dureté…
FIGURE 3-1
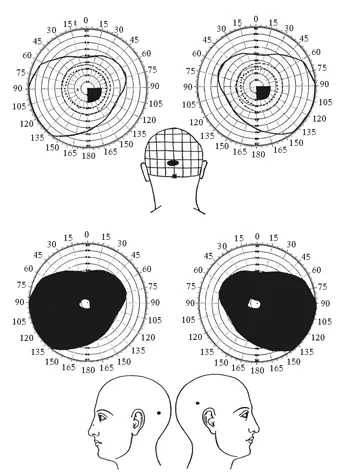
Représentation du champ visuel chez deux sujets présentant des blessures par balle de la région postérieure du cerveau. En haut, la lésion affecte la partie la plus postérieure et médiane du lobe occipital gauche. Les champs visuels de l’œil droit et de l’œil gauche présentent une zone aveugle (un scotome, représenté par la zone noire) situé dans la partie droite du champ visuel. Noter que ce scotome affecte seulement la vision de la partie centrale du champ visuel. En bas, la balle a traversé le crâne de part en part au niveau de la partie antérieure des deux lobes occipitaux. Le champ visuel des deux yeux présente un immense scotome qui n’épargne que la région centrale du champ visuel. Ces effets de lésions sur le champ visuel correspondent précisément à la topographie de la représentation du champ visuel sur le cortex occipital. Se reporter à la figure 3-2 pour l’anatomie de cette région.
Ces travaux résultent d’observations faites par le neurologue anglais Gordon Holmes sur des blessés de la guerre de 1914-1918.
Nous insisterons ici sur la manière dont le cerveau utilise les informations qui lui sont transmises de la surface du corps pour reconstruire le monde environnant et lui donner son sens. Prenons l’exemple de la surface sensible qu’est la rétine. La rétine, sur laquelle s’imprime l’image du champ visuel grâce aux lentilles de la partie antérieure de l’œil, envoie ses fibres vers une région spécialisée du cortex cérébral, le cortex visuel. À ce niveau existe une correspondance point par point entre une zone de l’espace visuel, une zone de la rétine et une zone du cortex. Cette correspondance (la rétinotopie) est due au fait que, comme nous l’avons vu, les fibres issues de la rétine se projettent sur le cortex de manière ordonnée, de telle sorte que leurs terminaisons y reconstituent une carte de la rétine. Ce mode d’organisation se retrouve dans tous les systèmes visuels connus de l’échelle des vertébrés possédant un cortex cérébral. Chez l’être humain, on a pu reconstituer cette carte à partir de l’observation de blessés de guerre porteurs de plaies du cerveau causées par des éclats d’obus. Une lésion localisée du cortex occipital entraîne une perte de la vision d’une région de l’espace (un scotome) (Figure 3-1). En cumulant les données obtenues chez de nombreux patients, on voit apparaître l’organisation topographique détaillée du cortex visuel : on peut ainsi repérer la zone correspondant à la fovéa (le centre de la rétine) ou la zone correspondant à son méridien vertical. Ce que nous montre cette reconstruction, c’est que la projection de la fovéa occupe sur le cortex une surface disproportionnée par rapport à sa surface réelle et à sa projection dans l’espace visuel. La vision au centre de la rétine a, de ce fait, une caractéristique remarquable : l’acuité visuelle y est sensiblement plus élevée que dans les autres parties de la rétine. Cette acuité élevée est la conséquence d’une densité plus forte en récepteurs rétiniens (les cônes). La partie centrale de la rétine bénéficie donc d’un facteur d’agrandissement plus élevé que les autres parties. Cela ne signifie évidemment pas que nous voyons plus gros les objets que nous fixons avec la fovéa : nous les voyons mieux, en y décelant des détails plus fins. En termes anatomiques, cela signifie que la vision de la fovéa dispose sur le cortex d’un plus grand nombre de connexions et donc d’un plus vaste espace de traitement (Figure 3-2).
FIGURE 3-2
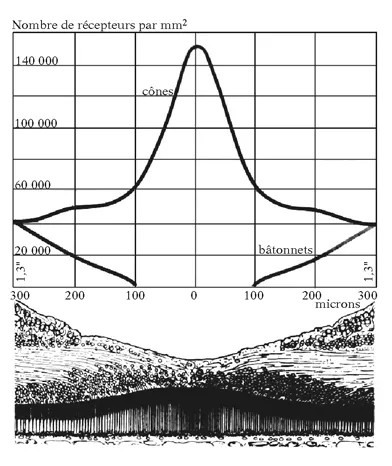
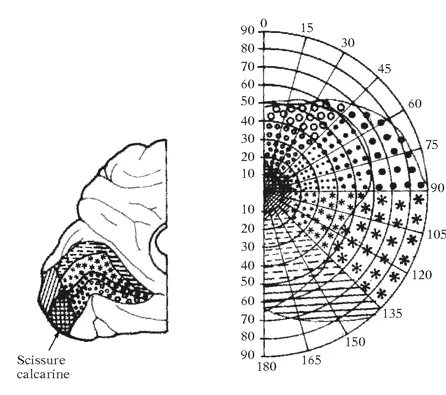
Caractère non linéaire de la rétinotopie sur le cortex occipital. Sur la page de gauche est représentée une coupe verticale de la rétine traversant la zone centrale (fovéa). Comme le montre le diagramme situé au-dessus de la coupe, cette zone est marquée par une forte densité en récepteurs (exclusivement des cônes), alors que les bâtonnets, moins nombreux, occupent la partie périphérique de la rétine. Ci-dessus est représentée la projection de la rétine (rétinotopie) sur la scissure calcarine du lobe occipital. On constate que la partie centrale de la rétine occupe à elle seule près d’un tiers de la projection de la rétine sur le cortex. Il existe donc un facteur de multiplication qui donne une plus grande surface synaptique à cette région fonctionnellement importante de la rétine.
La conséquence de cet arrangement anatomique est que ce ne sont pas les coordonnées réelles du monde extérieur qui sont représentées dans le cerveau : la carte rétinotopique est représentée en coordonnées fonctionnelles qui nous révèlent la façon dont les informations du monde extérieur sont traitées par le cerveau et l’utilisation que l’organisme fait de ces informations. En d’autres termes, la rétinotopie déformée par rapport à l’espace visuel que nous constatons sur le cortex nous révèle la lecture que le système nerveux fait du monde visuel. Pour prendre une comparaison géographique, on pourrait comparer la rétinotopie à la carte d’un pays où chaque région occuperait une surface qui serait fonction, non de sa taille réelle, mais de facteurs non topographiques, comme sa population, par exemple. De la même façon, la carte rétinotopique du cortex visuel représente les zones de la rétine en fonction de leur acuité. Chez l’être humain, le développement d’une acuité visuelle élevée dans la fovéa a nécessité une expansion de sa représentation corticale et permis l’apparition de nouvelles capacités visuelles. La lecture du langage écrit est une des nombreuses exploitations de ces nouvelles capacités.
Ce principe d’une organisation topographique du cortex cérébral non proportionnelle à la surface du récepteur, que nous venons de découvrir à propos de la rétine, se retrouve aussi pour d’autres récepteurs. La surface de la peau est représentée dans une région du lobe pariétal située près du sillon central (l’aire somesthésique), en face de la région motrice du lobe frontal. Là encore, la projection de la surface des différentes régions cutanées sur le cortex n’est pas proportionnelle à leur surface réelle. Les zones corticales correspondant à la projection des récepteurs sensoriels de la face et de la main occupent une part considérable de la surface du cortex, au détriment de vastes régions comme le thorax ou la jambe, par exemple, qui sont comparativement sous-représentées (Figure 3-3 à gauche).
Cette disproportion correspond à la finesse de la discrimination tactile au niveau de la face et surtout du bout des doigts qui nous servent à identifier les objets, leur texture, et le matériau dont ils sont faits. La face est remarquablement efficace pour la détection fine de la température d’un objet : on approche souvent un objet de sa figure pour estimer sa température.
La projection de la surface des récepteurs sensoriels et de l’ensemble de la surface du corps sur le cortex cérébral constitue une image de l’environnement sur le cortex cérébral. Cette image, toutefois, ne reproduit pas l’espace réel : c’est une image transformée, une reconstruction adaptée aux besoins de l’organisme. On ne comprend mieux l’importance de cette organisation topographique transformée que lorsqu’on examine ses relations avec le système moteur, celui qui permet la réalisation des mouvements. Pour qu’un mouvement dirigé vers un objet soit efficace, le système moteur doit posséder une structure qui corresponde le plus étroitement possible à la façon dont est traitée l’information sensorielle concernant cet objet. Le système moteur doit posséder une organisation topographique qui soit en concordance avec la représentation de l’espace visuel et de l’espace du corps. Le caractère non linéaire de cette représentation, qui pouvait nous paraître surprenant, devient ainsi une des conditions d’efficacité du transfert de la perception en action.
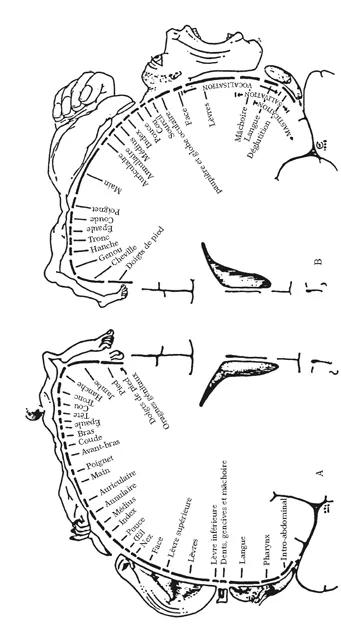
Représentation de la sensibilité et de la motricité sur le cortex cérébral. La figure de gauche montre la distribution de la sensibilité de la surface de la peau sur une coupe verticale du cortex passant par la région pariétale. Noter l’importance de la surface dévolue à la face, à la bouche (lèvres, pharynx, langue) et, dans une certaine mesure, à la main. Le reste du corps occupe une surface comparativement beaucoup plus faible. À droite est représentée sur une coupe passant par la région motrice du cortex moteur la distribution des régions contrôlant les muscles du squelette. Notez la surreprésentation des muscles de la face et de la main. Ces cartes ont été obtenues par le neurochirurgien canadien W. Penfield à partir de stimulations électriques du cortex de patients au cours d’opérations.
Une partie de l’information sensorielle traitée par le cerveau est en effet utilisée pour produire des mouvements, soit en réponse à une stimulation, soit spontanés et dirigés vers un but situé à l’extérieur du corps. Une partie importante du cerveau (la région motrice du cortex, comme nous l’avons vu, mais aussi de nombreuses autres régions comme le cervelet* et les corps striés*) est consacrée à la production et au contrôle de nos mouvements. Le cerveau moteur communique avec la musculature par l’intermédiaire de la moelle épinière : c’est là que se trouvent les motoneurones, cellules nerveuses qui entrent en contact direct avec les muscles. Les motoneurones reçoivent leurs instructions du cerveau par des faisceaux de fibres plus ou moins directs. Le plus important, chez l’humain, est le faisceau pyramidal qui, lui, provient directement de la région motrice du cortex cérébral. Chez les mammifères qui se sont développés antérieurement à l’apparition des primates, le faisceau pyramidal n’atteint qu’une faible partie des motoneurones. Chez le lapin, par exemple, il ne descend pas dans la moelle au-dessous des segments du cou, c’est-à-dire, n’innerve que les motoneurones des membres antérieurs. On peut expliquer ainsi la surprise des premiers expérimentateurs qui avaient constaté que la lésion du cortex moteur chez de tels animaux ne provoquait qu’une paralysie de courte durée : ils constataient qu’un animal sans cortex pouvait avoir un comportement moteur normal. Chez l’être humain, en revanche, la lésion du cortex moteur d’un côté ou la destruction d’un faisceau pyramidal entraîne une paralysie complète et persistante de la musculature de la moitié du corps (l’hémiplégie).
La représentation de la musculature à la surface du cortex moteur obéit à une organisation topographique du même type que celle de la surface de la peau. Les régions du cortex moteur qui innervent les muscles de la face et de la main sont surreprésentées par rapport à la masse réelle de ces muscles. Il en est de même des petits muscles du tractus vocal. À l’inverse, la commande des muscles du cou, du tronc et des membres inférieurs (à l’exception du pied) ne dispose que d’une surface corticale minime par rapport à leur masse. La carte motrice constitue donc une image déformée du corps, sous la forme d’un petit homme avec une tête et des mains de taille disproportionnée (Figure 3-3). Cette disproportion correspond à la finesse et à la précision des mouvements effectués à l’aide des muscles de la face (les mimiques et les expressions faciales), des muscles des doigts ou des muscles vocaux. Pour la face et les doigts, la finesse des mouvements est à l’échelle de la discrimination tactile. Au cours de la manipulation d’un objet, les mouvements de chaque doigt sont contrôlés indépendamment de ceux des autres, ce qui permet l’exploration de la forme et de la texture de l’objet par les récepteurs cutanés de la pulpe des doigts, là où les récepteurs tactiles sont les plus nombreux et les mieux représentés dans le cortex somesthésique. En outre, lorsque la manipulation se déroule sous le contrôle de la vision, les informations tactiles et visuelles coopèrent pour l’exploration de l’objet : au cours de cette opération, l’objet tenu par la main est placé dans la zone de vision correspondant à la zone centrale de la rétine, là où la vision est la plus fine. Nous disposons ainsi, dans notre espace de préhension, d’une capacité particulière adaptée à une reconnaissance plurimodale des objets (nous avons vu plus haut un autre exemple de reconnaissance plurimodale pour identifier un aliment dans la bouche). Cette capacité est le fruit d’une adaptation unique, propre aux primates et surtout à l’homme, qui nous permet de manipuler, de transformer et d’assembler des objets, en un mot d’utiliser des outils pour agir sur notre environnement.