
eBook - ePub
Learning and Memory
The Behavioral and Biological Substrates
Isidore Gormezano, Edward A. Wasserman, Isidore Gormezano, Edward A. Wasserman
This is a test
Share book
- 424 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Learning and Memory
The Behavioral and Biological Substrates
Isidore Gormezano, Edward A. Wasserman, Isidore Gormezano, Edward A. Wasserman
Book details
Book preview
Table of contents
Citations
About This Book
This volume presents the views and findings of behaviorally and biologically oriented investigators invited to participate in The University of Iowa's biennial learning and memory symposium. While chapters vary in their scope and depth of coverage, they are all amply referenced so that researchers, teachers, and students can obtain background information appropriate to their respective needs.
Frequently asked questions
How do I cancel my subscription?
Can/how do I download books?
At the moment all of our mobile-responsive ePub books are available to download via the app. Most of our PDFs are also available to download and we're working on making the final remaining ones downloadable now. Learn more here.
What is the difference between the pricing plans?
Both plans give you full access to the library and all of Perlego’s features. The only differences are the price and subscription period: With the annual plan you’ll save around 30% compared to 12 months on the monthly plan.
What is Perlego?
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 1000+ topics, we’ve got you covered! Learn more here.
Do you support text-to-speech?
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more here.
Is Learning and Memory an online PDF/ePUB?
Yes, you can access Learning and Memory by Isidore Gormezano, Edward A. Wasserman, Isidore Gormezano, Edward A. Wasserman in PDF and/or ePUB format, as well as other popular books in Psychology & History & Theory in Psychology. We have over one million books available in our catalogue for you to explore.
Information
1 | Foraging by Rats on a Radial Maze: Learning, Memory, and Decision Rules |
The research I discuss represents a blending of two areas of investigation, spatial memory and foraging. Beginning with the seminal article of Olton and Samuelson (1976), a number of experiments have been carried out on the radial maze to find out how rats and other animals remember and forget spatial information. This research has revealed that rats can remember a large number of locations visited (Olton, Collison, & Werz, 1977; Roberts, 1979) and that the format of memory is maplike and based on extramaze visual cues (Mazmanian & Roberts, 1983; Suzuki, Augerinos, & Black, 1980). Although rats can remember locations visited on the radial maze over several hours (Beatty & Shavalia, 1980), forgetting can arise from both proactive and retroactive interference caused by other spatial experiences (Roberts, 1981; Roberts & Dale, 1981).
If one reflects on the functional role of spatial memory, it seems clear that spatial memory promotes fitness in rats and other animals (Shettleworth, 1983). Specifically, spatial memory allows animals to remember where in space a number of places are located that are vital to survival, such as food locations, the location of home base, and the locations of potential predators and conspecifics. Spatial memory also allows an organism to keep track of its own movements through a spatial landscape and prevents redundant visits to locations recently visited. Finally, animals not only remember locations in their environment but also remember the contents of those locations or important events that occurred at those locations.
Much of this concern with the functional role of spatial memory is incorporated within the second area of investigation, foraging theory. Spatial memory may be seen to be at the service of foraging strategies that tend to maximize the survival of an organism or its genes. Thus, spatial knowledge informs decisions that tend to maximize energy gained over time by a forager and its offspring and to minimize exposure to predation.
Shettleworth (1989) has recently stressed the distinction between functional and mechanistic foraging questions. Many models of foraging advanced by biologists have addressed functional questions about the way in which evolutionary processes would shape optimal foraging behavior. Psychologists, on the other hand, have tended to focus on proximal questions about the more immediate mechanisms that govern foraging decisions in existing animals. Presumably, these two approaches to the study of foraging go hand in hand; that is, mechanisms of foraging evolved to serve functions that tend to optimize foraging behavior. For example, if it is optimal for a forager to visit environmental locations containing food and to avoid ones not containing food, there should be evolutionary pressures for the selection of organisms that can learn and remember food locations.
Much of the foraging work in psychology has been carried out in Skinner boxes, with animals working for reinforcements on concurrent and multiple schedules that deliver reinforcement at different rates on a random or quasi-random basis. The question usually asked in such experiments is whether an animal will learn to switch between keys or schedules at times that tend to maximize the overall energy gain (e.g., Bhatt & Wasserman, 1987; Dow & Lea, 1987; Fantino & Abarca, 1985; Kamil, Yoerg, & Clements, 1988). Relatively few experiments have been carried out in the psychology laboratory to investigate what may be a relatively more realistic situation of animals foraging across a spatial field in which food may be found in different locations. Further, few experiments have been done to study foraging when the forager may have complete or nearly complete knowledge of an environment and its contents.
Within natural settings or seminatural environments constructed in the laboratory, evidence of the important functional use of spatial memory in foraging has begun to accumulate. Menzel (1973, 1978) allowed chimpanzees to observe as many as 18 food items being hidden at different locations in an open outdoor field. When released, the chimpanzees foraged in a least-distance-traveled pattern and thus minimized the time and effort required to accumulate all the available food. When confronted with an abundance of food that cannot be consumed at the moment, some species of birds hoard available food and recover it at a later time. In both the field and the laboratory, it has been shown that Clark’s nutcrackers can remember a vast number of locations where pinyon pine seeds have been buried days before (Balda, 1980; Balda & Turek, 1984; Kamil & Balda, 1985). Marsh tits and chickadees allowed to hide food in a natural setting or in holes in trees in a laboratory show foraging efficiency by selectively returning to those places where food was hidden (Sherry, 1987; Shettleworth & Krebs, 1982, 1986). By using spatial memory for the location and contents of food sites, foraging birds save valuable time and energy that would be lost if a search not informed by memory were used.
A few recent laboratory studies with rats suggest foraging efficiency. Mellgren and his associates examined the foraging patterns of rats confronted with food items buried in boxes of sand that were dispersed about a large room (Mellgren, 1982; Mellgren & Brown, 1987; Mellgren, Misasi, & Brown, 1984). Rats searched longer in patches as patch density and travel time between patches increased and showed the most efficient foraging when patch density was unpredictable and patches were difficult to reach. Several radial maze studies suggest that rats are very sensitive to the contents of maze arms and tend to prioritize their visits to arms on the basis of their contents. Hulse and O’Leary (1982) placed different quantities of reward pellets in food cups at the ends of the arms of a four-arm radial maze and found that rats came to enter the arms with the largest amounts before the arms with the smaller amounts. In other experiments, water-deprived rats have encountered water or other liquid substances placed on the arms of a radial maze. Rats learned to avoid an arm that did not contain water and to prefer one that contained chocolate milk (Batson, Best, Phillips, Patel, & Gilleland, 1986). When consumption of saccharin was followed by lithium chloride poisoning, rats avoided any arm of a radial maze that contained saccharin (Batson et al., 1986; Melcer & Timberlake, 1985).
In the experiments reported here, I show that rats foraging on a radial maze are sensitive to several variables that are directly relevant to foraging in a natural setting. These variables include structural aspects of the foraging situation, such as differences in density of food among patches, location of food within a patch, travel time between patches, and ease of access to food placed in feeders, and properties of food items, such as number and size. In several cases, rats’ behaviors will appear illogical or unnecessary within the laboratory context. However, consideration of this behavior from an evolutionary point of view suggests that it has definite survival value. It is argued that the radial maze has considerable ecological validity for the study of foraging in rats, and that this ecological validity may be a result of the radial maze mimicking to a certain extent the burrow system of wild rats.
Optimal Foraging by Rats
In several experiments my students and I have carried out recently, we have extended the use of the radial maze for studying foraging in rats by adding certain constraints on foraging. In Fig. 1.1, a top diagram is shown of a four-arm radial maze used in my laboratory. Notice that each arm contains four feeding stations or food cups attached to a platform on the side of the arm. In the experiment depicted in the figure, the arms of the maze were defined as patches, and the patches contained 0 (0% patch), 1 (25% patch), 2 (50% patch), or 3 (75% patch) baited feeders. The baited feeders (F) each contained a pellet of General Mill’s Count Chocula breakfast cereal. Information was varied in this experiment. One group of rats was allowed to forage with the food locations remaining fixed throughout 24 foraging sessions. This group then had the opportunity to learn food locations and to make use of reference memory to forage efficiently. A second group of rats had food locations varied randomly from one session to the next; the four patches always contained 0, 1, 2, or 3 baited feeders, but patch density and the specific locations of baited feeders within patches varied randomly between daily sessions. Two other constraints on foraging were manipulated. Travel time between patches was manipulated by requiring rats to climb over wooden barriers placed at the entrance to each arm. On alternate daily sessions, barriers were either present or absent. As a third constraint, the time and effort required to gain access to feeders was raised by requiring rats to push metal covers off the feeders to get at their contents. One subgroup of rats within each of the fixed and random food location conditions was tested with covered feeders, and another subgroup was tested with open feeders.
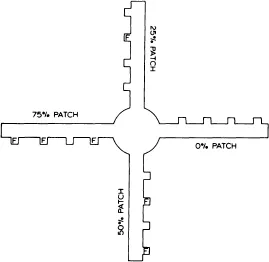
Fig. 1.1. Diagram of a four-arm radial maze with four feeding stations on each arm (patch). Patch density was varied by placing food (F) in 0 (0% patch), 1 (25% patch), 2 (50% patch), or 3 (75% patch) of the feeders in each patch. (From Roberts & Ilersich, 1989. Copyright 1989 by American Psychological Association. Reprinted by permission.)
In Fig. 1.2, two fixed food location curves and two random food location curves are shown for animals foraging with barriers placed at patch entrances and covers placed over feeders. One curve represents optimal foraging as determined by a computer simulation. The simulations were done by using average times for different components of foraging taken from the final sessions of testing (see Roberts & Ilersich, 1989) and computing the times at which food items 1-6 would be consumed if an optimal strategy were used. In the case of fixed food locations, the optimal strategy was entering patches in decreasing order of density and visiting only baited feeders within each patch. With random food locations, the optimal strategy was to enter four different patches and exhaustively visit all four feeders within the patch. The observed curves represent the actual times at which rats consumed items on the final sessions of testing. Note that the fixed food location group collected all food items substantially sooner than the random food location group. An important observation to be made is that both groups’ curves are only slightly below the optimal foraging curves. The observed points were generally within 10% of the times generated for the optimal curves. In other words, rats were making foraging decisions that kept their rate of food accumulation near optimality.
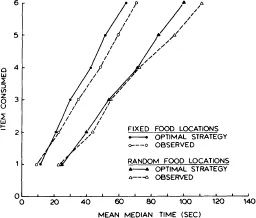
Fig. 1.2. Gain functions showing mean median times at which successive food items were consumed. The optimal strategy curves are based on computer simulations, and the observed curves are based on rats foraging for food in fixed or random locations on a four-arm maze.
Foraging Decision Rules
We may inquire about the mechanisms or decision rules rats used in this experiment to stay near optimal foraging. All three of the constraining variables manipulated had an effect on foraging decisions.
The Effect of Travel Time
When travel time between patches was raised by placing barriers at patch entrances, rats in the random food location condition stayed in patches longer and visited all the feeders in the patch before leaving. The rate of patch reentry then was low. Without barriers, rats stayed for a shorter time in patches, less frequently visited all the feeders within a patch, and more frequently revisited patches. These findings suggest that rats were willing to abandon patches without exhaustively searching them when the price of a revisit was low, but not when the price of a revisit was high.
Discrimination Among Patches
The difference between fixed and random observed curves seen in Fig. 1.2 suggests that rats were using reference memory to locate food items sooner in the fixed food location group. Evidence for food location learning can be sought at two levels of choice decisions: choice among patches varying in food density and choice between baited and unbaited feeders within patches. Discrimination among patches is examined in Fig. 1.3, in which the mean rank of entry into patches varying in food density is plotted across blocks of two daily sessions. Lower ranks indicate early patch entrance, and higher ranks indicate late patch entrance. The data from the random food location group are shown in the left panel and indicate that rats never discriminated among patches. This finding is important because it indicates that rats could not detect patch density by the use of odor cues. In the right panel, rats developed a strong tendency to enter the 0% patch late in the sequence of patch visits. In many cases, rats simply did not enter the 0% patch. Although there is some average tendency for rats to enter the 75% patch before the 50% and 25% patches, the differences among these three patches were not statistically significant. These findings indicate that rats strongly discriminated between patches with food and a patch without food. The finding that they did not discriminate strongly among patches with different numbers of baited feeders suggests that decisions among these patches might have been based on a decision process in which an entry decision was made whenever a scan of reference memory yielded at least one baited feeder.
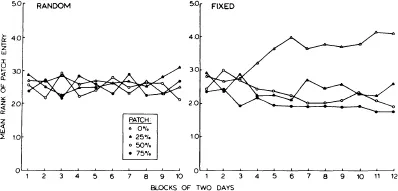
Fig. 1.3. Mean ranks of first entry into patches varying in density plotted over blocks of 2 days. Data for the random food locations condition are shown in the left panel, and data for the fixed food locations condition are shown in the right panel. (From Roberts & llersich, 1989. Copyright 1989 by American Psychological Association. Reprinted by permission.)
Discrimination Among Feeders
Rats tested with fixed food locations came to discriminate well between baited and unbaited feeders within pat...