
- 300 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Evolution of the Brain: Creation of the Self
About this book
Sir John Eccles, a distinguished scientist and Nobel Prize winner who has devoted his scientific life to the study of the mammalian brain, tells the story of how we came to be, not only as animals at the end of the hominid evolutionary line, but also as human persons possessed of reflective consciousness.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
Mind & Body in PhilosophyIndex
Biological SciencesChapter one
Biological evolution
1.1 The genetic code
In order to be able to present an intelligible story of the essentials of the evolutionary process, it is necessary first to give a much-simplified account of the genetic material of the cell, deoxyribonucleic acid (DNA), and of the mode of its action via the genetic code. The segregation of this essential evolutionary material into the cell nuclei was achieved very early in the evolution of the unicellular eukaryotes that arose about 1.8 billion years ago. This was a very important evolutionary development because it protected the complex machinery that is central to all cell activity including reproduction.
The DNA of the nucleus is a densely coiled and extremely long double helix. As diagrammed in Figure 1.1, each strand is constructed of alternate phosphate (P) and sugar (ribose) moieties. To each sugar there is attached one of the following four molecules: the purine bases, adenine (A) and guanine (G), and the pyrimidines, thymine (T) and cytosine (C). The two helices are effectively cross-linked every 3.4 Å (Figure 1.1). A in one links to T in the other or G in one to C of the other. So a sequence could be:
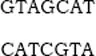
for the linkage pairs of a very short segment of the two helices. The nucleotide code is thus written linearly along each strand. Figure 1.2a shows the atomic structure of the double helix with below the phosphate (P) and sugar (S) chain and the cross-linkage by the purine and pyrimidine bases through the hydrogen bonding of A with T and C with G. Figure 1.2b illustrates the manner in which the linear code of the DNA is translated to messenger RNA (ribonucleic acid), which effects the segmental building of the amino acid sequences of a protein by means of a three-letter code acting like a machine language in specifying the sequence of the five amino acids in this specimen record.
For a bacterium the code of each strand has about 1.5 million letters. With Homo there are about 3.5 billion letters in each DNA strand, which gives the preliminary information for building all of the cells of a human being. Before the cell divides, the two strands of the double helix separate and an enzyme system makes for each the complementary strand. The two double helices that are thus reconstituted are almost always identical copies of the original. The genetic information that builds and controls the cell is coded in the nucleotide sequences, the ATGC letters, along the DNA strands.
It is beyond the scope of this chapter to go into the detailed manner in which, by the precise processes of transcription and translation, this DNA code is read out in the building of the amino acid sequences of a protein (see Figure 1.2b), and so is effective in building the structure of the cell and in the enzymatically controlled metabolism of the cell. Enzymes are proteins. The code for any such action in building a protein is carried in linear array on the DNA strands, not by a short sequence of letters as illustrated above, but by some thousands of letter sequences, called a gene. Genes carry the precise instructions for building the amino acid sequences of particular proteins. It will be recognized that, for building the many species of proteins required for the living processes of a bacterial cell, its DNA chain of about 3 million letter sequences is not extravagant. With our cells the number is more than 1,000 times greater, 3.5 billion. This seems rather extravagant for coding the information required to build the proteins of our cells. It has been estimated by Dobzhansky (personal communication) that the number of human genes is at least 30,000. For an average protein of 500 amino acid sequences, 1,500 nucleotide pairs are required, because three pairs are required for each sequence. So 30,000 genes require 4.5× 107 nucleotide pairs. However, with redundancies the number could be many times larger, so lifting this low ratio of 1.4 per cent.
Moreover there is the unsolved problem of the ‘silence’ of at least 30-70 per cent of the mammalian genome. A partial solution is in the DNA spacers, which are sequences separating the active DNA segments.
Normally in reproduction there is an accurate copying of the linear code written in the DNA, and hence there is stability in the genes from generation to generation. However, changes called gene mutations do occur in the DNA code. There may be mistakes in copying with the replacement of one nucleotide for another, such as G for A, or there may be more radical changes with deletion or inversion of one or more nucleotide base pairs or even inversion of larger DNA segments. These copying errors may lead to the substitution of one amino acid for another in a protein. The effect of this may be negligible in the functioning of the protein. However, in the great majority these exchanges are deleterious to the survival and reproduction of the individual, which consequently is eliminated in the process of natural selection.
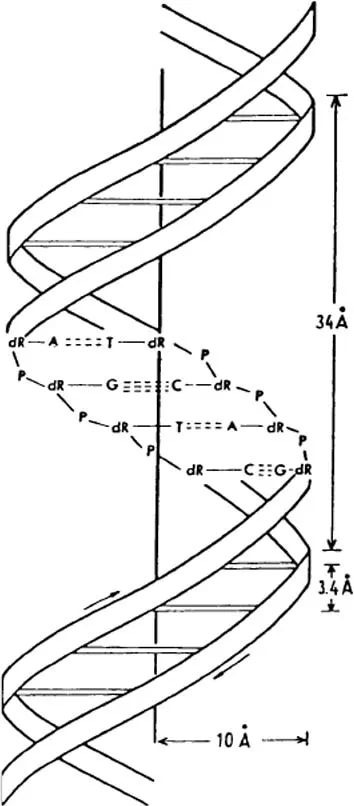
Figure 1.1 The double-stranded helical configuration of the DNA molecule. The two nucleotide strands are held together by hydrogen bonds forming between the complementary purine (A or G) and pyrimidine (T or C) pairs. Note the dimensions given for the spacing, and for width and length of one helical configuration.
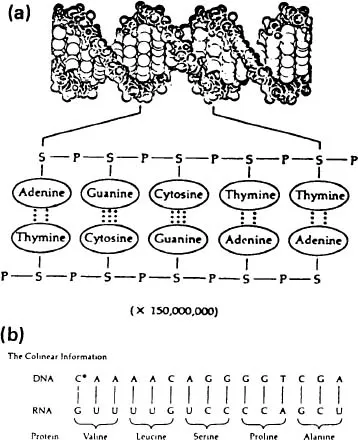
Figure 1.2 (a) Above: atomic structure of DNA molecule. Below: diagram of connections in DNA-S, sugar, P, phosphate, covalent bonds as lines, hydrogen bonds as dots. (b) Diagram of a small segment of DNA with the processes of transcription to RNA and translation from RNA to amino acids. From Darwin to DNA, Molecules to Humanity by G.Ledyard Stebbins Copyright © WH Freeman and Co. Reprinted with permission.
Only on rare occasions is a mutation beneficial for survival and reproduction. Such a mutation will be transmitted to successive generations and will result in enhanced survival of the biological group sharing this mutation. So after many generations by natural selection this favourable mutation may come to be incorporated in all members of that species, which consequently reflect a slight change in genotype. Later another mutational selection may be added, and so on.
This is the essential basis of the modern version of Darwin's theory of natural selection or survival of the fittest. Favourable gene mutations are selected, whereas the unfavourable are eliminated.
Hence by an initial process of pure chance, the gene mutation, there can be wrought by natural selection all the marvellous structural and functional features of living organisms with their amazing adaptiveness and inventiveness. As so formulated, the evolutionary theory is purely a biological process involving mechanisms of operations that are now well understood in principle, and it has deservedly won acceptance as providing a satisfactory explanation of the development of all living forms from some single extremely simple primordial form of life. This theory, stemming from Darwin and Wallace, must rank as one of the grandest conceptual achievements of man. Yet it is in need of remodelling (Sections 1.2 and 1.3 below).
A recent development has been the recognition that many mistakes in DNA copying are virtually neutral. For example, the mutation may result in a changed amino acid sequence in a part of the protein that is not vital for its functioning. Or, again, the mutation may be in a part of the DNA that is not concerned in building protein, so it will be selectively neutral. In time there can be a large accumulation of such neutral mutations that have changed considerably the original DNA of a population. With a changed environment these mutations may no longer be neutral.
The DNA of a cell nucleus does not exist as an enormously long double helix of about 2 m in length, but is subdivided into segments that compose the chromosomes, which become evident when the cell is in the process of meiosis during subdivision. Then the human genome is seen to be packaged into 23 pairs of chromosomes each with its distinctive character (Figure 1.3). In meiosis the chromosomes with their contained DNA subdivide and separate to form the sex cells, and, when there is fertilization, the full complement of DNA is reconstituted, half coming from each sex cell.
The four living species of Hominoidea (Table 2.1) are very similar in their nuclear structure. The three species of pongids-chimpanzees, gorilla, and orangutan-have 48 chromosomes. In Homo two pairs of chromosomes have united by centric fusion to form chromosome 2, hence Homo has 46 chromosomes (Figure 1.3). In other respects there is a remarkable similarity, even to the details of the banding patterns along the chromosomes of the respective species.
1.2 The modern synthesis: phyletic gradualism (Mayr, 1963)
Ever since Darwin it has been recognized that biological species play the key role as units in evolution. A species consists of a population rather than of unconnected individuals. The population of a species is reproductively isolated from all other species because of the fertility criterion. Other rather similar species may inhabit the same territory, but despite this sympatric coexistence there is no interbreeding. ‘Each’ species is a delicately integrated genetic system that has been selected through many generations to fit into a definite niche in its environment' (Mayr, 1963:109).

Figure 1.3 A photograph of a normal complement of chromosomes of a human female enlarged 15,000 times. The normal number of chromosomes in the human is 46. (Handler, 1968.)
In the Darwinian perspective palaeontology accounted for the formation of new species by transformation of the ancestral population by a very slow process with large numbers of individuals in the inhabited territory. It is a process that Eldredge and Gould (1972) called phyletic gradualism. Unfortunately this gradualness is not shown in the fossil record. The classical evolutionists attribute this deficiency to the imperfection of the fossil record. The fossils indicate a story of sharp breaks or saltations in the evolutionary process. The genetic diversity of a species is due to mutations, recombinations, deletions, etc. in the genetic transmission from one generation to the next. However, it is controlled by the collective process of gene flow in the successive generations of a freely breeding population. Nevertheless, no two individuals of a sexually reproducing population are genetically identical (Mayr, 1963), except for identical twins.
Despite the homogenizing effect of gene flow, the modern synthetic theory of phyletic gradualism ‘continued this tradition of extrapolation from local populations and used the accepted model for adaptive geographical variation-gradual allelic substitution directed by natural selection-as a paradigm for the origin of species’ (Gould, 1982:134; ‘alleles’ is used here as a collective name for genes). This global concept has been called sympatric continuity. However, Mayr (1963) recognized that speciation could occur more rapidly and effectively in small isolated populations. A small founding population would migrate so that it is isolated from the gene flow of the large ancestral population. But this model adhered to the principle of phyletic gradualism in the peripheral isolate. The successful speciation occurred because of the cumulative effects of small adaptive variations through a large number of generations. The advantage of the isolate was solely due to the diminished homogenizing effect of the gene flow within the small population.
1.3 Punctuated equilibrium
Eldredge and Gould developed a theory of allopatric speciation in which a new species can arise
only when a small local population becomes isolated at the margin of the geographic range of its parent species. Such local populations are termed peripheral isolates. A peripheral isolate develops into a new species if isolating mechanisms evolve that will prevent the re-initiation of gene flow if the new form re-encounters its ancestors at some future time. As a consequence of the allopatric theory, new fossil species do not originate in the place where their ancestors lived. (1972: 94)
Eldredge and Gould (1972) postulate that the development of a new species in the peripheral isolate occurs in a short period of time relative to the duration of the species, and, if there is migration back to the territory of the ancestral species, the two species will coexist sympatrically without interbreeding. This can be observed in the fossil record. Thus long periods of stasis are punctuated by episodic events of allopatric speciation. This is the hypothesis of punctuated equilibria.
As mentioned above, phyletic gradualism depends on unitary gene mutations, which, if adaptive and successive, are gradually accumulated by selection over long periods of time. There are now alternative models that give fast punctuational effects. Major chromosomal changes may give the genetic changes requisite for speciation in a few generations of a peripheral isolate. Thus speciation may be dependent on gene regulation and rearrangement rather than on the classical point mutations that produce new genes in phyletic gradualism. Also large phenotypic changes may result from changes in the timing of regulatory genes, which in this way would cause the production of a new species (Bush et al., 1977). Carson proposed that:
speciational events may be set in motion and important genetic saltations towards species formation accomplished by a series of catastrophic, stochastic genetic events…initiated when an unusual forced reorganization of the epistatic supergenes of the closed variability system occurs… I propose that this cycle of disorganization and reorganization be viewed as the essence of the speciation process. (1975:88)
This proposed saltatory origin of species is not adaptive as is the case in classical phyletic gradualism with selection as the key control of the random point mutations. Reproductive isolation with the large fast saltatory genetic changes comes first and is not adaptive. Gould (1982) goes so far as to argue that, though the saltatory formation of species provides the raw material for selection, there is a diametric difference between these two alternative theories of speciation. According to phyletic gradualism, point mutations lead to allelic substitutions in local populations that are sequential, slow and adaptive by selection. According to the punctuational equilibrium hypothesis, the saltatory origin of new species is discontinuous and nonadapting and is only secondarily subject to selection.
There may be saltatory production of an extreme character producing what is ironically termed a hopeful monster. Selection may determine whether or not a ‘hopeful monst...
Table of contents
- Cover page
- Title page
- Copyright page
- Foreword by Sir Karl Popper
- Preface
- Acknowledgements
- List of abbreviations
- Chapter one: Biological evolution
- Chapter two: The general story of human evolution
- Chapter three: Evolution of hominid brain: bipedality; agility
- Chapter four: Linguistic communication in hominid evolution
- Chapter five: Cerebral limbic system in relation to the evolution of the reproductive and emotional systems
- Chapter six: Visuo-motor evolution: artistic creativity
- Chapter seven: Evolution of learning and memory
- Chapter eight: The mind—brain problem in evolution
- Chapter nine: Creation of the self and its brain
- Chapter ten: The human person
- Addendum: Reflections and imaginings
- Abbreviations used in References
- References
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Evolution of the Brain: Creation of the Self by John C. Eccles in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Mind & Body in Philosophy. We have over 1.5 million books available in our catalogue for you to explore.