Plant Defense provides an overview of all major aspects of plant defence, including defence against pathogens, parasites, and invertebrate and vertebrate herbivores. The book looks at defense mechanisms including structural and chemical defences, and constitutive and inducible defences. Including details of how plants 'sense' attack and how this is communicated within the plant and also to neighbouring plants, how plants coordinate defence responses to simultaneous multiple attacks, and the energy and resources expended by a plant in maintaining and implementing its defence systems.

eBook - ePub
Plant Defense
Warding off attack by pathogens, herbivores and parasitic plants
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Why Do Plants Need Defenses?
1.1 Plants as sources of food
All organisms need food to survive. To be more precise, they require a variety of chemical elements—the most important of which are carbon, nitrogen, and oxygen—to provide the building blocks for growth and development. This in turn requires a supply of energy, the only external supply of which comes from the sun. Plants are able to capture the energy from sunlight and convert it into chemical energy, thereby providing the means of financing the formation of carbohydrate from atmospheric CO2 and water. This autotrophic ability of plants comes at a price. Because most organisms are not autotrophic, they must obtain their energy and building blocks for growth and development from consuming other organisms, including plants. In fact, plants are a direct source of food for an array of organisms that include invertebrates, vertebrates, fungi, bacteria, and even other plants.
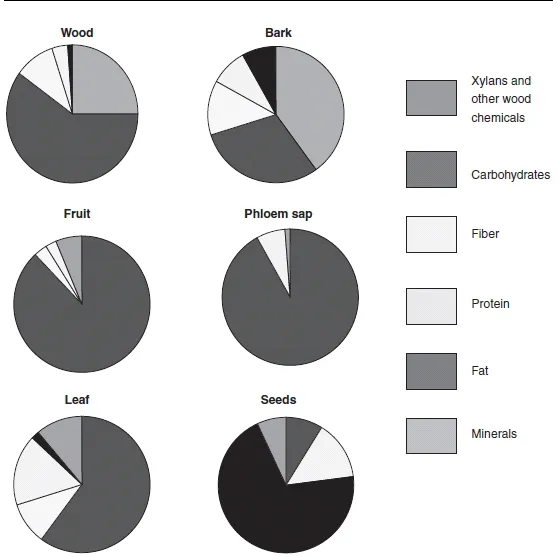
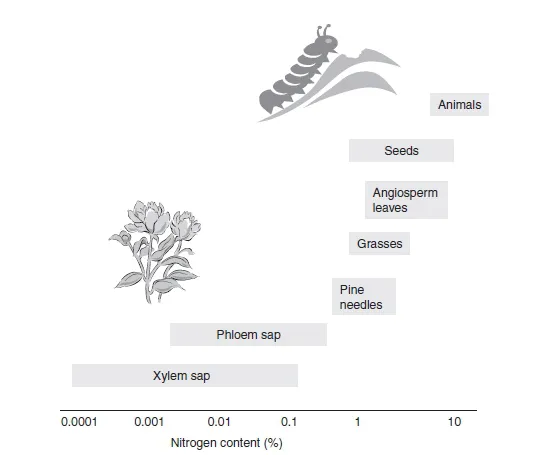
The popularity of plants as food sources for so many organisms begs the question “what do plants offer other organisms by way of nutrition?” Clearly, plant tissues will provide a source of carbon and nitrogen, much of which will be in the form of carbohydrates, lipids, andproteins.Theywillalsocontainmacroelementssuchasphosphorus,sulfur,calcium,and potassium, as well as various microelements such as iron, manganese, and zinc. However, the relative proportions of these components will vary depending on species. Moreover, different plant parts can have very different compositions (Figure 1.1). For example, fruits and phloem sap can be rich sources of carbohydrates, while seeds are usually good sources of fat. Some parts of the plant, such as bark, offer little in the way of nutrients, since they are composed largely of dead cells, with lignified walls. Nitrogen and protein content also varies between different parts of the plant, but in general, plants contain less nitrogen and protein than most of the organisms that use them as a food source. Typically, the total nitrogen content of plants is between 2 and 4% of their dry weight, while the nitrogen content of animals amounts to 8-14% of their dry bodyweight (Figure 1.2). The amount of nitrogen in insects is even greater than this and can be in the order of 30-40% of their dry weight (Southwood, 1973).
As indicated above, plants are used as food sources by a variety of organisms. Before we proceed further, it is worth considering the mechanisms used by these organisms to obtain the nutrients locked up in plant tissues.
Figure 1.1 Composition of different plant parts that can serve as food for herbivores. (Adapted from Townsend et al. (2003), with permission of Blackwell Publishing Ltd.)
1.2 Organisms that use plants as food
1.2.1 Microorganisms
Plants are infected by a wide range of microorganisms. Some of these establish symbiotic associations with plant roots, such as bacteria found in nodules on roots of legumes and mycorrhizal fungi, which form intimate associations with the roots of most plant species. Other microorganisms are parasitic on plants and use plants as food sources, causing damage and sometimes plant death, in the process. Some of these microbes, including viruses, protozoa, and some fungi, are biotrophs. These grow and reproduce in nature only on living hosts. Powdery mildew and rust fungi (Figure 1.3a), for example, produce feeding structures called haustoria that invaginate the host plasma membrane, forming an intimate association with the plant cell. Other microbes, mostly fungi and bacteria, are necrotrophic. They secrete enzymes to cause disintegration of plant cells and, either alone or in combination with toxins, often lead to cell and tissue death (Figure 1.3b). The nutrients released in the process are then available for uptake by the pathogen.
Figure 1.2 Variations in nitrogen concentration (dry weight percentage) of different plant parts compared with that in animals. Xylem and phloem sap concentrations are expressed as nitrogen weight/volume. (Adapted from Mattson (1980), with permission.)
1.2.2 Parasitic angiosperms
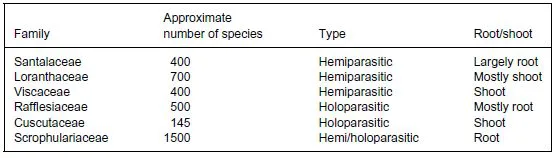
Plants are also parasitized by other plants. Indeed, parasitism among plants seems to have evolved many times during angiosperm evolution. It has been estimated that about 1% of angiosperms, some 3000 species in total, are parasitic on other plants (Parker & Riches, 1993). Parasitic angiosperms are distributed among 17 families, including the Viscaceae and the Cuscutaceae (Table 1.1), and include parasitic plants such as species of and (Figure 1.3c and 1.3d). There is considerable diversity in the extent to which parasitic angiosperms rely on the host for growth. Some, such as species of, have functional roots and can therefore take up inorganic nutrients from the soil, while others, such as the mistletoes, have nothing that resembles a root nor functions as one (Hibberd & Jeschke, 2001). There is also considerable variation in the extent to which parasitic plants rely on the host for photoassimilates. Thus, parasitic plants such as Rhinanthus minor are able to photosynthesize and can grow with a carbon supply from the host, while others, such as, possess a very low photosynthetic capacity and are unable to grow without a carbohydrate supply from the host (Hibberd & Jeschke, 2001).
Figure 1.3 (a) Yellow rust (Puccinia striiformis) on wheat, (b) Alternaria brassicae on pods and stems of oilseed rape (Brassica napus), (c) the parasitic angiosperm Rhinanthus minor, (d) the parasitic angiosperm Orobanche crenata, (e) the plant parasitic nematode Helicotylenchus, (f) caterpillar of the large cabbage white butterfly, Pieris brassicae, (g) an aphid on a leaf, (h) moose, (i) cows grazing. (Image (c) is the copyright of Glyn Baker and is licensed for reuse under the Creative Commons Licence. Images (d), (e), (f) and (h) are reproduced courtesy of Lytton Musselman, the American Phytopathological Society, Rosemary Collier of the University of Warwick HRI, and the United States Geological Survey, respectively.)

Table 1.1 Main families of parasitic plants
1.2.3 Nematodes
Nematodes are wormlike in appearance (Figure 1.3e) but are quite distinct from the true worms. Several hundred species are known to feed on living plants, obtaining their food with spears or stylets. Nematode feeding causes only slight mechanical damage to plants. The majority of the damage caused by nematodes appears to be caused by saliva injected into plants during feeding. Nematodes will puncture a cell wall, inject saliva into the cell, and withdraw part of the cell contents. Some nematodes feed rapidly and move on within a few seconds, while others feed more slowly and remain at the puncture for hours or even days. As long as the nematodes are feeding, they will inject saliva intermittently into the cell.
1.2.4 Insects
There are more species of insects than any other class of organisms on earth and nearly half of these, some 400,000 species, feed on plants (Schoonhoven et al., 2005). These herbivorous insects harvest their food inavariety of different ways. Chewing insects possess “toothed” mandibles that cut, crush, and macerate plant tissues. Many feed externally on the plant, while others, such as leaf miners, harvest tissue layers between the upper and lower epidermis of the leaf. Chewing insects include species belonging to the orders Lepidoptera (moths and butterflies) (Figure 1.3f) and Orthoptera (e.g., grasshoppers). Some insects do not remove chunks of plant tissue, but rather suck fluids from the plant using specialized tubular mouthparts. Thus, insects in the order Hemiptera include aphids (Figure 1.3g), which feed on phloem sap. Other members of the Hemiptera, for example, whiteflies, feed on the contents of leaf mesophyll cells. Yet other insects make galls on their plant host. These insects manipulate the host tissues, providing themselves with both shelter and nutrients. A striking feature of relationships between insects and plants is the extent of food specialization among insect herbivores. Some insects, including many lepidopterous larvae, hemipterans, and coleopterans, occur on only one or a few closely related plant species and are termed monophagous. Others, such as the Colorado potato beetle,, are oligophagous; these feed on a number of plant species, all belonging to the same family. Yet other insect herbivores, for example, the aphid, accept many plants belonging to different families. Such insects are polyphagous.
Table 1.2 Major categories of dietary specialization in herbivorous mammals, according to Eisenberg (1981)
| Category | Diet |
| Nectarivores | Nectar and pollen |
| Gumivores | Exudates from trees |
| Frugivore/omnivore | Pericarp or fleshy outer covering of plant reproductive parts, invertebrates and small vertebrates |
| Frugivore/granivore | Reproductive parts of plants, including seeds |
| Frugivore/herbivore | Fleshy fruiting bodies and seeds of plants, storage roots, and some green leafy material |
| Herbivore/browser | Stems, twigs, buds, and leaves |
| Herbivore/grazer | Grasses |
1.2.5 Vertebrates
Herbivory is not confined to insects. In fact, it is a common trait among mammals (Figure 1.3h and 1.3i), with roughly half of the 1000 or so genera of mammals including plants in their diet (Danell & Bergström, 2002). While the majority of herbivorous insects are mono- or oligophagous, feeding on a few plant species, vertebrate herbivores tend to be polyphagous and feed on a wider range of plant species. Vertebrate herbivores are larger than their invertebrate counterparts and are thus able to remove a greater amount of plant tissue with each mouthful (Danell & Bergström, 2002). The classification of animals into functional groups is usually achieved using diet composition. Sixteen major categories of dietary specialization in mammals have been proposed (Eisenberg, 1981), and of these, seven refer to herbivores (Table 1.2). Categories range from nectarivores that feed on nectar and pollen to gumivores that feed on exudates from trees. The most dominant group is numerically the frugivores/omnivores, which represent approximately 33% of vertebrate herbivore genera (Table 1.2).
1.3 Impact of infection and herbivory in natural andagricultural ecosystems
1.3.1 Microorganisms
Pathogenic microorganisms can exert a profound effect on the structure and dynamics of individual plant species and plant communities. The extent and type of damage to individual plants is related to the lifestyle of the pathogen, that is, whether it is a biotroph or a necrotroph. Necrotrophs destroy plant tissue and it seems obvious therefore, that loss of leaf tissue, for example, will decrease rates of photosynthesis, thereby reducing plant growth. In contrast, biotrophs do not kill plant tissue, although effects on photosynthesis can be just as profound. Thus, effects on chloroplast structure and function can lead to dramatic reductions in rates of photosynthesis in plants infected with rust or powdery mildew fungi (Walters & McRoberts, 2006). Photosynthesis can also be affected by other means. In plants infected with vascular wilt pathogens, such as Verticillium albo-atrum, blockage of xylem vessels can lead to water stress and partial closure of stomata, thus reducing rates of photosynthesis. Of course, the effects of pathogens on the host plant are not restricted to photosynthesis, and some pathogens, for example, can alter water and nutrient uptake, while others produce toxins, which affect host metabolism. Whatever the mechanism, pathogen infection can lead to greatly reduced plant growth and reproductive output. Thus, Albugo candida and Peronospora parasitica, both biotrophs, reduce reproductive output in Capsella bursa-pastoris (Alexander & Burdon, 1984), while the tobacco leaf curl virus reduces growth and seed production in its host, Eupatorium chinense, and is an important cause of plant mortality (Yahara & Oyama, 1993).
Pathogen infection can also lead to plant death. Thus, fungal damping off and root diseases can cause mass mortality of seedlings, especially under humid conditions. Damping off was responsible for 64-95% of seedling deaths of the tropical tree Platypodium elegans in the first 3 months after emergence (Augspurger, 1983). Pathogens can also cause death of older plants. In Australia, an epidemic of the root rot pathogen, Phytophthora cinnamomi, devasted a dry sclerophyll forest (Weste & Ashton, 1994), while the pathogen Phacidium infestans was a significant cause of mortality in 5- to 10-year-old Pinus sylvestris (Burdon et al., 1994).
In crop production systems, losses due to pathogens can be substantial. In the period 1996-1998, global crop losses due to pathogens (fungi, bacteria, and viruses) were 12.6%, in spite of crop protection measures (Oerke & Dehne, 2004). Some crops seem to suffer more than others, and between 1996 and 1998, pathogens accounted for losses of 22% in global potato production. Even more devastating can be the spread of a pathogen into a new geographical area. For example, the soybean rust, Phakopsora pachyrhizi, was first reported in South America in 2001 (in Paraguay) and by 2003 was detected in most soybean-growing regions of Brazil, with losses estimated at 5% of total soybean production (Yorinori et al., 2005).
1.3.2 Parasitic angiosperms
The diversion of host resources to parasitic plants can have large effects on host growth and reproductive output. Infection of Poa alpina with the annual hemiparasite, R. minor, reduced host biomass by more than 50% (Seel & Press, 1996), while the phloem-tapping mistletoe, Tristerix aphyllus, greatly reduced the production of buds, flowers, and fruits by its cactus host, Echinopsis chilensis (Silva & Martínez del Rio, 1996). Death of the host plant can occur, particularly in extreme cases, such as heavy infestation with mistletoe (Aukema, 2003).
Parasitic plants can exert a considerable impact on plant communities (Press & Phoenix, 2005). Thus, Rhinanthus species have been shown to reduce total productivity in European grasslands by between 8 and 73% (Davies et al., 1997), while dwarf mistletoes can reduce volume growth of Douglas fir by up to 65% (Mathiasen et al., 1990).
Striga is a genus of root hemiparasite with some 35 species, most of which are of no agricultural importance. However, those species that parasitize crop plants can be devastating. Yield losses in cereals infected by Striga can reach 100%, and fields can be so heavily infested that they are abandoned by farmers (Berner et al., 1995). Some 40 million hectares of cereals are thought to be severely infested with Striga spp. in West Africa, and the Food and Agriculture Organization (FAO) estimates that annual yield losses in the savannah regions alone account for US$7 billion (Berner et al., 1995).
1.3.3 Nematodes
Nematode infestation can lead to substantial re...
Table of contents
- Cover
- Title page
- Copyright
- Dedication
- Preface
- Chapter 1: Why Do Plants Need Defenses?
- Chapter 2: What Defenses Do Plants Use?
- Chapter 3: Sounding the Alarm: Signaling and Communication in Plant Defense
- Chapter 4: Plant Defense in the Real World: Multiple Attackers and Beneficial Interactions
- Chapter 5: The Evolution of Plant Defense
- Chapter 6: Exploiting Plant Defense
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Plant Defense by Dale Walters in PDF and/or ePUB format, as well as other popular books in Technology & Engineering & Agriculture. We have over 1.5 million books available in our catalogue for you to explore.