![]()
CHAPTER 1
FROM ISOLATED ACTIONS TO TRUE MUSCLE FUNCTION
PAAVO V. KOMI
Neuromuscular Research Center, Department of Biology of Physical Activity, University of Jyväskylä, Jyväskylä, Finland
Introduction
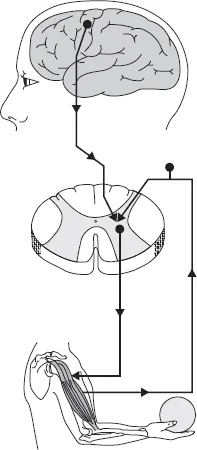
In order to function properly for force and power production in locomotion, the skeletal muscle possesses structural elements, which make it very responsive to commands from the nervous system, both centrally and peripherally. It is not intended to go into the details of the nervous control of the muscle contraction in this chapter. However, certain important aspects are needed to clarify the role the nervous system plays in muscle function and movement control. For more specifics, the reader is referred to general textbooks of neurophysiology and motor control. In the simplest form of force generation, the central nervous system (CNS) triggers the chain of activation along the direct pathway (pyramidal or corticospinal tract) as shown in Figure 1.1. The signal can also come from the different brainstem motor areas along the indirect pathways labeled as extrapyramidal tract. The activation signal from the motor areas passes then through several stages before reaching the muscle fibers to activate events which include Ca2+ release from the sarcoplasmic reticulum (SR) and subsequent contraction (excitation–contraction coupling). In addition, and very importantly in motor control, the signal can be initiated in the various muscle, joint, skin receptors, etc. The nervous system operates as a controller being able to initiate action potentials, receive and integrate feedbacks from the higher brain centers and from the spinal cord levels. Consequently, even when the activating signal travels along the simplest and most direct pathway (corticospinal tract, CST) it is under the influence of inputs from different sources and levels of the nervous system. All this information, regardless of the source, is received and integrated by the alpha motor neuron, which is called the “final common pathway of the motor system.” The type of muscle contraction is then determined by the frequency of action potentials coming from the alpha motor neuron. The alpha motor neuron can itself have different properties, as will be discussed shortly in connection with “fast” and “slow” motor units. Skeletal muscle contains all of the elements needed for force and movement production, but without the nervous control, the muscle is not capable of any force production above the passive tension. The passive tension (or resting tension) maybe due to the muscle’s structural elements, which offer resistance to stretch. Nonetheless, the muscle can be activated by impulses coming along the final common pathway, the alpha motor neuron. Upon activation, it then has a special ability to generate force, resulting in either shortening (concentric action) or resisting external loads (lengthening contraction or eccentric action). A complex integrative process involving the three components, the nervous system, skeletal muscle, and the external load, determines the final direction of movement as well as its velocity (or rate) and magnitude. It is the purpose of this chapter to characterize those factors that are important in understanding the basic interaction between the elements mentioned earlier. Greater emphasis will, however, be given to the important concepts of muscle mechanics as well as to the interaction between the contractile structures and tensile elements in the process of force production under varying movement conditions.
The Motor Unit and its Functional Significance
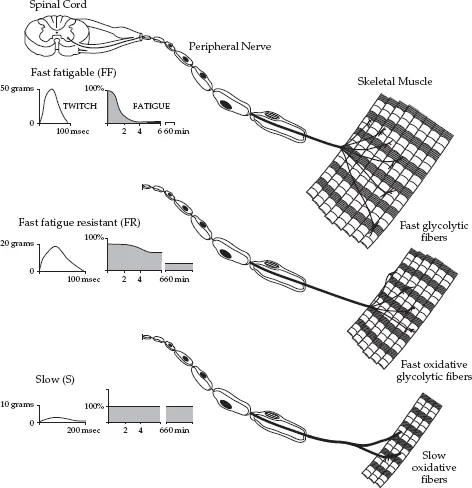
It is usually believed that human skeletal muscle fibers are innervated by only one motor neuron branch, but this branch maybe one from 10 to 1000 similar branches all having the same axon. Therefore, one axon innervates a number of muscle fibers and this functional unit is called a “motor unit.” Consequently, a motor unit is defined as a combination of an alpha motor neuron and all the muscle fibers innervated by that neuron. Motor unit size (muscle fibers per alpha motor neuron) varies within a muscle, and the number of motor units varies between muscles. As illustrated in Figure 1.2, the motor units have different structural and functional characteristics, which result in their differences with regard to rate of force development (RFD), peak force production, and maintenance of force level without loss of tension (fatigue). The fast, fatigable (FF) type unit develops tension quickly, but is also very easily fatigued. At the opposite end, the S-unit has a slow rate of force production but can produce the same tension (force) repeatedly for longer periods of time without signs of fatigue. It is therefore also called a “fatigue-resistant motor unit.” In addition to those events described in Figure 1.2, there are also other functional differences between motor unit types. One particular feature that illustrates such differences is the response of the motor units to tetanic stimulation The FF type unit requires a high stimulation frequency to reach a state of tetanus. In contrast, the slower unit requires a much lower fusion frequency. When subjected to repetitive tetanic stimulation, the resulting difference in mechanical response between the two extreme types of motor units is remarkable. It is most probably the size of the alpha motor neuron that determines the specific type of the motor unit. Motor neurons supplying the faster motor units have larger cell bodies and axons. They also innervate larger number of individual muscle fibers. One important feature must be emphasized here: the type of alpha motor neuron determines the histochemical profile and biochemical performance of the individual muscle fibers in a motor unit. Consequently, all the fibers in the same unit have similar chemical profile. It is well known in the literature that muscles differ in their fiber composition (and thus in their motor unit profiles), and that there can be great variation among athletes with regard to the fiber structure in a specific muscle. For example, in the vastus lateralis (VL) muscle, sprinters may have a motor unit make up that causes most of the fibers in that muscle to be of a fast type, and thus capable of producing force at a high rate, but with low fatigue resistance. Endurance runners, on the other hand, have primarily slow type fibers in the same muscle for the purpose of high resistance to fatigue, but at the same time the rate of force production is lower than in their sprinter counterparts. It has been reported from studies with monozygotic twins that genetic factors strongly influence the variation observed among individuals in muscle fiber composition (Komi et al., 1977) of a specific muscle. Differences in muscle fiber composition observed among athletes have thus raised the question as to whether the fiber composition of an individual athlete is an acquired phenomenon or is due to a genetically determined code.
As discussed in more detail in Chapter 13, the order of the motor unit recruitment follows the so-called “size principle” (Henneman, 1957). The influence of size on recruitment order is attributable to its effect on input resistance. The small motor neurons have a high input resistance, and they are the first to be recruited in response to an increase in depolarizing synaptic currents. As a consequence, smaller motor units are activated before larger units. Due to the relation between the size of the motor neuron and the properties of the muscle fibers it innervates, this recruitment sequence results in slow-contracting and fatigue-resistant motor units being recruited before fast-contracting and fatigable motor units. Although there is some variability in the recruitment order of motor units with similar thresholds, the recruitment order of motor units is essentially the same for isometric and dynamic contractions, including shortening and lengthening contractions and during rapid (ballistic) isometric and shortening contractions.
Muscle–Tendon Mechanics
Skeletal muscle has different properties, which influence its functional characteristics. Based on the earlier discussion about the different types of motor units (e.g., fast versus slow), the property of the alpha motor neurons should then be mostly responsible for the final product of the function: mechanical performance of the muscle. The human skeletal muscle is not, however, only the muscle; it contains an important element of the tendon, which in its turn has special mechanical characteristics. Consequently, the performance of skeletal muscle is under the influence of the innervation, the histochemical make ups of the contractile elements and the tendon, which connects the muscle to the insertion sites on the bones. In human (or animal at large) body, these elements and profiles must function together to produce a well-controlled movement. In addition, a muscle in the body acts often together with its synergist and antagonist muscle. The final mechanical output is therefore a complex phenomenon, and it is almost an impossible task to predict the true movement from measurements performed with individual components (activation profile, contractile part, and tendon) only in an isolated form. However, the contribution of the scientists, who have done pioneering work with isolated preparations, must not be forgotten. This work, which started already in the early 1900s, has been fundamental to understanding muscle mechanics in vivo locomotion. Several steps needed to be taken and the following paragraphs make attempts to present this basic information.
Twitch Characteristics of Isolated Muscles
As already referred to, action potentials traveling along the final common pathway (alpha motor neuron) reach finally the t-tubule and release calcium from its stores in the SR. Even a single action potential results in calcium release with subsequent binding with the troponin C and generation of force in the actomyosin cross-bridges. Almost at the same time the free calcium is taken back into SR. During this short-lived “active” period when calcium is attached to troponin C, the generated force can be recorded, and it is called “twitch.” The rate of rise of the twitch tension as well as its relaxation are under influence of several factors, including the availability of free calcium, the speed of the binding of the myosin heads, rate of the calcium uptake back to SR, and the rate of the cross-bridge dissociation. Both the rates of the force development and relaxation are under influence of elastic properties of the cross-bridge. Most importantly, however, it is the activation profile that determines the twitch properties of the motor unit or the entire muscle.
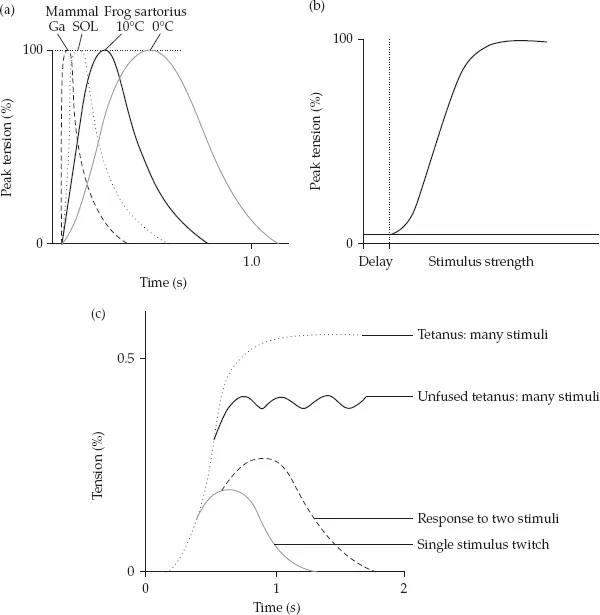
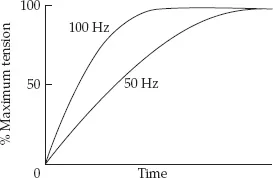
When the electrical stimulation is used for twitch experiments, its strength must be strong enough to depolarize the muscle fiber (or muscle) membrane. Single stimuli of identical strength should then produce exactly similar force records (twitches). This indeed happens, if the two stimuli are separated by suitable interval. However, when the second stimulus is given before the first twitch is over, the resulting second force peak is usually higher than the first one (see Figure 1.3). The increase of the compound force becomes more pronounced the closer the consecutive twitches are brought together. This is illustrated in Figure 1.3. If the time interval between twitches is reduced so that the train of shocks comes with the frequency of 30 Hz, the force records of the consecutive twitches do not return to zero. If the stimulation frequency is further increased, e.g., to 50 Hz, the compound record may still increase and the curve looks like an unfused tetanus. Further increase in stimulating frequency (until 100 Hz, corresponding to a 10 ms stimulus interval) will finally result in complete fusion of the twitch responses. Thus as the frequency is increased, the ripple over the force record is reduced. Relationship between degree of this oscillation and the mean (or sometimes peak) force is used to imply the speed of muscle. Fast twitch muscle usually has a higher tetanic frequency as compared to the slower muscle. In a particular muscle, however, the 50 Hz stimulation is usually enough to reach almost the full maximum isometric force level, and the additional increase of frequency up to 100 Hz does not necessarily increase the peak force, but has considerable influence to increase the RFD (see Figure 1.4). This force–frequency relationship has given the basis for the force–time (F–T) relationships in the conditions of maximal voluntary activation. The fast type muscle can increase the force much faster than the slow type muscle, and this is primarily due to the faster rate of activation. This has subsequently been applied to strength and power training, in which the increase in the RFD indicates improvement in the explosive force production. As explained in Chapter 13, specificity of training calls for modification of the F–T curve, the changes of which take place in the different parts of the curve depending on the training intensities and loads.
There are also indications that young girls have much slower rates of force development as compared to their male counterparts of the same age category (Komi et al., 1977). This was interpreted to indicate that females may have slower activation profile as the boys in the explosive F–T test, where the force production is started with zero activation. This difference may not be so apparent in more normal locomotion, such as stretch-shortening cycle (SSC) type muscle function, in which the preactivity plays an important role in efficient force and power production (see Chapter 2). Further discussion of the importance of the F–T curve and its practical relevance is given in the following paragraphs.
Basic Muscle Mechanics
Types of Muscle Action
In order to understand the way that skeletal muscle functions in normal locomotion, the relation between stimulus and response needs to be examined in more isolated forms of muscle actions: isometric, concentric, and eccentric. The term “contraction” maybe thought of as the state of muscle when it is activated via its alpha motor neurons, and generates tension across a number of actin and myosin filaments. Depending on the external load, the direction and magnitude of action is different as shown in Table 1.1. In a concentric action, the muscle shortens (i.e., the net muscle moment is in the same direction as the change in joint angle and mechanical work is positive). In an eccentric action, the muscle actively resists while it is being lengthened by some external force, such as gravity. In this case, the resulting muscle moment is in the opposite direction to the change in joint angle, and the mechanical work is negative. The use of the term “muscle contraction” is therefore sometimes confusing, and we would prefer to follow a suggestion made by Cavanagh (1988) that the term “contraction” should be replaced by “action.”
Table 1.1 Classification of muscle action of exercise types
| Concentric | Acceleration | Positive (W F(D)) |
| Isometric | Fixation | Zero (no change in length) |
| Eccentric | Deceleration | Negative (W F(D)) |
The muscle action most frequently used to characterize the performance of human skeletal muscle is the isometric action, which by definition refers to the activation of muscle (force production) while the length of the entire muscle–tendon unit (MTU) remains the same, and the mechanical work is zero The use of isometric action in locomotion is not, however, meaningless; it plays a very important role in the process of preactivation of the muscle before the other actions take place.
Force production in all types of muscle actions can be seen in the internal rearrangements in length between t...