
- 640 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Research Methods for Cognitive Neuroscience
About this book
This fresh, new textbook provides a thorough and student-friendly guide to the different techniques used in cognitive neuroscience. Given the breadth of neuroimaging techniques available today, this text is invaluable, serving as an approachable text for students, researchers, and writers. This text provides the right level of detail for those who wish to understand the basics of neuroimaging and also provides more advanced material in order to learn further about particular techniques.
With a conversational, student-friendly writing style, Aaron Newman introduces the key principles of neuroimaging techniques, the relevant theory and the recent changes in the field.
Tools to learn more effectively

Saving Books

Keyword Search

Annotating Text

Listen to it instead
Information
1 The Organization of the Brain and How We Study It
Learning Objectives
After reading this chapter, you should be able to:
- Define the scope and objectives of the field of cognitive neuroscience.
- Identify the fundamental types of cells in the brain, and how they communicate with each other.
- Describe the basic organization of the brain, across micro-, meso-, and macro-anatomical scales.
- Explain the value of measuring behaviour in understanding both cognition and brain activity.
- List the various brain imaging and stimulation methodologies covered in this book, and categorize them based on the type of data they measure.
Introduction
Cognitive neuroscience is the field of study aimed at understanding how the brain produces thoughts, emotions, and behaviour. By and large, this field focuses on human beings specifically, and – given the general reluctance that most humans express to having their heads cut open – relies primarily on non-invasive methods for characterizing brain activity and structure. The field of cognitive neuroscience is quite new; although humans have a long-standing interest in the thoughts and behaviours of themselves and others, the idea that ‘There could be a human neurobiology of normal cognitive processes’ (Gazzaniga, 2018) was realized only three decades ago. Specifically, in 1988 a group of researchers from Washington University and the University of Oregon published the first studies of human cognition using positron emission tomography (PET) – a form of brain imaging that allowed researchers to localize changes in blood flow in the brain, using radioactively labelled oxygen (Petersen, Fox, Posner, Mintun, & Raichle, 1988; Posner, Petersen, Fox, & Raichle, 1988). Prior to this, the only ways to study brain activity in healthy, living humans employed EEG, which involves measuring brain electrical activity via electrodes attached to the outside of the head (or a related technique, MEG; however, this was restricted to a very small number of labs). Although as we will see in this book EEG is an extremely valuable tool, it does not provide accurate information as to where in the brain activity originates, and so left many unanswered questions. Other tools existed for studying brain–behaviour relationships, but these necessarily involved unhealthy brains, such as neuropsychological studies of people with developmental disorders or acquired brain damage, and direct electrical recordings from the surface of the brain made during neurosurgery.
A few scant years after the publication of the first PET studies, a number of other techniques were established for studying brain activity non-invasively, including functional MRI (fMRI) and near-infrared optical imaging (fNIRI), as well as transcranial magnetic stimulation (TMS) for non-invasively and transiently perturbing brain function. These all contributed to the development of the new field of cognitive neuroscience, which was codified with the establishment of a dedicated, peer-reviewed scientific journal – the Journal of Cognitive Neuroscience – in 1989, and the first meeting of the Cognitive Neuroscience Society in San Francisco in 1993. These milestones were, however, the fruition of work going back decades earlier by trailblazers who were willing to ask the hard questions, and seek answers even within the extremely limiting constraints of existing technologies. Perhaps most importantly, these early investigators were willing to transcend the traditional disciplinary boundaries of cognitive psychology on the one hand, and neurophysiology and neuroanatomy on the other. As Michael Gazzaniga, founding editor of the Journal of Cognitive Neuroscience, wrote in the Editor’s Note to its inaugural issue,
Those cognitive scientists interested in a deeper understanding of how the human mind works now believe that it is maximally fruitful to propose models of cognitive processes that can be assessed in neurobiologic terms. Likewise, it is no longer useful for neuroscientists to propose brain mechanisms underlying psychological processes without actually coming to grips with the complexities of psychological processes involved in any particular mental capacity being examined. (Gazzaniga, 1989: 2)
This book focuses on these and other techniques that have revolutionized our understanding of the human brain, and the nature of human thoughts, feelings, and actions. These techniques have fascinated me for many years, dating back to when I was in high school and saw the pioneering PET research mentioned above reported in Discover magazine. In a fit of characteristic nerdiness, I remember bringing that issue of the magazine to school to show my friends, and my disappointment that they were not nearly as excited about it as I. Nevertheless, my interest persisted and I had the opportunity to first work with functional MRI during my undergraduate degree in 1995–96, thanks to the enthusiasm of my supervisor, Dr Michael McIntyre, and the National Research Council of Canada. Mike in turn connected me with Dr Helen Neville at the University of Oregon, who accepted me as a graduate student and provided me with amazing opportunities to learn EEG and fMRI, and connect with pioneering researchers from around the world. Among the many things I learned from Helen were that, as amazing as the tools of cognitive neuroscience are, they are only small windows into the workings of the brain, and are highly fallible. In consequence, it is vital to always question your data, inspect it closely (and repeatedly) for errors, and remember that results are only as good as the experimental design, and even then always very much subject to interpretation. Simply obtaining a result does not make it true.
From Cells to Networks
Although our interest in cognitive neuroscience is in linking the brain with cognition and behaviour, as a first step it is critical to have an understanding of the organ we are studying and, in particular, of the levels of organization of the brain that we are able to study with the tools of cognitive neuroscience. This book will be most understandable if the reader has some prior background in both psychology and neuroscience; however, in this section we will review the bare essentials. This is intentionally a very simplified account and readers are encouraged to consult an introductory neuroscience textbook for a more detailed explanation.
Structural Units
The adult human brain weighs approximately 1.5 kg and is composed of roughly 86 billion neurons (nerve cells), and a roughly equal amount of non-neuronal cells (Herculano-Houzel, 2009; Purves et al., 2017). Neurons are the key players in the transmission of information throughout the brain and the body, forming synapses with other neurons so that electrical signals can be transmitted from one neuron to another. The non-neuronal cells are predominantly glia, which were traditionally viewed as ‘helper’ cells supporting neuronal functions (such as modulating activity), but which in recent years have come to be appreciated as important functional units in the brain as well (Magaki, Williams, & Vinters, 2017; Verkhratsky & Kirchhoff, 2007) and, as we will see, are also critical to generating the signal we measure with fMRI.
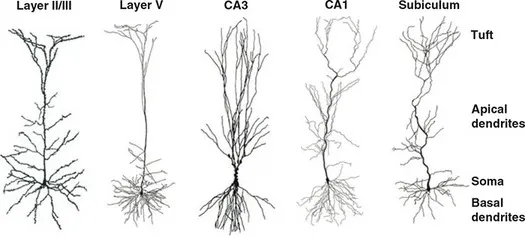
Neurons are, however, the main actors in cognitive function. There are many different types of neurons in the brain. One dominant type of neuron found in the cerebral cortex of humans (as well as most other mammals, birds, fish, and reptiles) is the pyramidal cell. Pyramidal cells are found primarily in brain structures supporting higher cognitive functions, including the cerebral cortex, amygdala, and hippocampus (Spruston, 2008), making them of particular relevance for cognitive neuroscience – although all types of neurons are no doubt important for brain function and cognition. Nevertheless, we will focus on pyramidal cells here to exemplify neuronal structure and function. As illustrated in Figure 1.1, pyramidal cells typically have a cell body or soma, from which extends a single axon that branches extensively, sometimes along its entire length and in other cases only at the end (the tuft). These branches are called dendrites, and these are where the majority of connections (both input and output) are made with other neurons. Dendrites also branch out from the soma, with the apical dendrites being those along the axon, and basal dendrites branching directly off the soma. Functionally distinct roles have been identified for different regions of pyramidal cells at an even finer-grained level, with several distinct subregions of both the apical and basal dendritic regions (Spruston, 2008).
Figure 1.1 Pyramidal neurons from different parts of the brain, including layers of the cerebral cortex (leftmost two images; indicated by Roman numerals) and the hippocampus (rightmost three images). Labels on the right side indicate different regions of the cells. Reprinted from Spruston, 2008 with permission of Springer Nature
Neurons are densely connected with other neurons, over both short and long distances. It is these patterns of synaptic connections that allow the brain to carry out its complex representations and computations. Neurons communicate by way of both electrical and chemical transmission. At rest, neurons are electrically polarized, meaning that the electrical potential within the cell is negative relative to the space around the cell. Electrical potentials are covered in detail in Chapter 2, but for now we can simply say that there is potential for electrical charge to move between the outside and inside of the neurons (in the form of ions such as sodium, potassium, and calcium). A primary way for neurons to transmit information is to ‘fire’, or generate an action potential, in which case ion channels on the neuronal membrane open and allow the electrical charge inside the neuron to equilibrate with that outside the neuron – a phenomenon known as depolarization. This change in electrical potential starts at the soma and propagates down the axon, ultimately resulting in the neuron’s firing being communicated to other neurons it is connected to. Importantly, axons are typically covered in a fatty coating called myelin, which serves as electrical insulation to both increase the speed of electrical transmission, and prevent the signal’s strength from weakening along the length of the axon. The myelin sheath is actually provided by a specific type of glial cell, called oligodendrocytes. Figure 1.2 shows an example of an action potential travelling from a neuronal cell body, down its axon to the terminal where it communicates with another neuron.
Figure 1.2 A schematic illustration of two neurons connected via a synapse
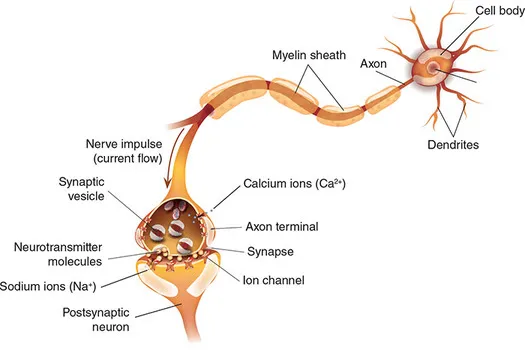
The connections between neurons are called synapses, but it is important to understand that neurons are not directly physically connected to each other, but rather are separated by gaps, as shown in Figure 1.2. Thus when an electrical signal reaches the end of the axon, it does not directly propagate to the connected neurons. Instead, an arriving action potential triggers the release of chemical messengers called neurotransmitters. These chemicals are released by presynaptic (‘sending’) neurons to signal postsynaptic (‘receiving’) neurons by fitting into receptors: molecular structures on the neuronal membranes (outer walls of the cells) that fit the molecular shape of the neurotransmitter analogous to how a key fits a particular lock. The primary neurotransmitter used by neurons to excite other neurons (an excitatory neurotransmitter) is glutamate. There are three primary types of glutamate receptors: N-methyl-D-aspartate (NMDA), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA), and kainate. Although all three receptor types are activated by glutamate, they differ in the types of cells and locations on cells that they occur, and their names reflect the fact that they can be selectively activated by specific chemicals other than glutamate – meaning that certain drugs can modulate the activity of one type of glutamate receptor with little or no effect on the other types. Using such drugs is a common way to investigate the role of particular neurotransmitters and receptors in brain function and cognition. Drugs that selectively target a particular type of receptor are called agonists for that receptor. In contrast, drugs that selectively block a receptor (meaning that the normal neurotransmitter will not activate that receptor) are called antagonists.
Not all neurotransmission is excitatory; many neurons actually inhibit other neurons, making them less likely to fire. The primary inhibitory neurotransmitter in the brain is gamma-aminobutyric acid, or GABA, for which there are two primary types of receptors (with much more logical names than the glutamate receptors): GABAA and GABAB. In addition, there are other classes of chemicals, called neuromodulators and neurohormones, that affect neuronal activity. These act like neurotransmitters in the sense that they modulate neuronal function via receptors on the neurons. They differ from neurotransmitters in that rather than serving to communicate a transient action potential from one neuron to another at a very small scale across a synaptic junction, they are in many cases released into the intracellular space and act to modulate the function of larger numbers of neurons, making them more or less likely to fire in response to other inputs. Neuromodulators and neurohormones also act over longer time scales; while neurotransmitters act specifically at local synaptic junctions and are typically reabsorbed very quickly either by the neurons, or by surrounding glia (which then ‘recycle’ the neurotransmitters and send them back to the neurons), neuromodulators and hormones have longer lifetimes in the intracellular space prior to absorption or breakdown. Neuromodulators include serotonin, dopamine, norepinephrine, and acetylcholine, although some of these also act as short-acting neurotransmitters in some cell types and brain regions. Neurohormones function similarly to neuromodulators, and indeed the lines between the two are somewhat blurry, with different authors sometimes using one or the other to refer to the same chemical (Peres & Valena, 2011). However, neurohormones tend to refer to chemicals that are neuromodulatory, but produced in organs other than the brain and then travel through the body to the brain to modulate neural activity. In addition to the neuromodulators listed above, neurohormones include oxytocin, oestrogen, testosterone, vasopressin, insulin, and cortisol.
Networks of Neurons
Given the billions of individual neurons in the brain, it is not surprising that they form extremely complex, interconnected networks. The number of neurons connected to (that is, forming synapses with) another neuron varies widely by type and location of the cells, but estimates of around 10,000 connections per neuron are common. Within the cerebral cortex – the part of the brain that plays a primary role in most cognition (see next section) – there are many distinct regions that can be defined by the types and relative densities of cells present, the types of connections they have, and many other parameters. Even within a brain region, the cerebral cortex can be divided into distinct layers based on the locations and types of cells and connections. Typically six layers are defined within the cerebral cortex, although this may be further broken into sub-layers. This complex, layered organization of the cortex was first documented by Ramón y Cajal, who received the Nobel Prize in Physiology or Medicine for his work in 1906 (shared by Camillo Golgi, who invented the staining technique that made Cajal’s work possible). Two of Cajal’s drawings illustrating the laminal (layered) structure of the cerebral cortex are shown in Figure 1.3. Some layers are dominated by long-range inputs from other brain regions, while other layers contain horizontal cells – neurons with short-range connections that serve primarily inhibitory purposes.
Figure 1.3 The laminar (layered) structure of the cerebral cortex, as illustrated by early anatomists. In all drawings, the outer surface of the cortex is at the top, and the bottom of the drawings is the white matter underlying the cortex. The top and bottom left drawings are by Spanish neuroanatomist Ramón y Cajal, using the Golgi staining method. The large cells are pyramidal neurons, arranged in a consistent manner with their...
Table of contents
- Cover
- Half Title
- Acknowledgements
- Title Page
- Copyright Page
- Acknowledgements
- Contents
- Preface
- Acknowledgements
- About the Author
- How to Use This Book
- 1 The Organization of the Brain and How We Study It
- 2 Research Methods and Experimental Design
- 3 Electroencephalography (EEG) and Event-Related Potentials (ERP) 1
- 4 Electroencephalography (EEG) and Event-Related Potentials (ERP) 2
- 5 Magnetoencephalography (MEG)
- 6 Magnetic Resonance Imaging (MRI)
- 7 Functional MRI (fMRI) 1
- 8 Functional MRI (fMRI) 2
- 9 Structural MRI
- 10 Connectomics: Diffusion Tensor Imaging (DTI) and Tractography
- 11 Positron Emission Tomography (PET)
- 12 Near-Infrared Optical Imaging (fNIRI)
- 13 Transcranial Magnetic Stimulation (TMS)
- 14 Transcranial Electrical Stimulation (tES: tDCS, tACS, tRNS)
- Glossary
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Research Methods for Cognitive Neuroscience by Aaron Newman in PDF and/or ePUB format, as well as other popular books in Psychology & Research & Methodology in Psychology. We have over one million books available in our catalogue for you to explore.