![]()
Chapter 1
Studying the distribution of life on Earth
A biogeographic hiatus ended in the mid-1970s in a flood of analytical and theoretical papers that heralded a new phase of biogeography. For much of the early 20th century, biogeography was thought of as a by-product of evolutionary biology – an artefact of past climatic processes and events that helped shape physiological adaptions and dispersal routes. That was shattered by the acceptance of continental drift that was once rejected as ‘absurd’.1 Continental drift, in the guise of plate tectonic theory, had revolutionised the geosciences, including palaeontology, but it had little overall effect on biologists and how they practised biogeography. Physiological adaptions, mostly to climate, and dispersal routes were all that 20th century biogeography had to offer. Descriptive biogeography (the study of biotic regions and biota) was considered outdated and not in the same league as ecology and population dynamics. Did biogeographers simply dismiss geological processes as occurring over long periods of evolutionary time – too long to have any effect on the distributions we see today? For example, the drying out of Australia over a period of 20 million years was always thought to be due to ‘climate’ and nothing more. Now we know that this aridity is due to tectonics and fluctuations in the mantle causing the Australian plate to buckle and bend as it slowly moves north. Plate tectonics had usurped climate as the main driver of environmental change. Without the hot molten core and mantle that drives plate tectonics, our planet would resemble Mars: a cold and dead dust bowl. Even by the end of the 20th century, many biogeographers were still struggling with the idea of Earth and life evolving together. But not all of biogeography ignores a dynamic Earth; rather it is how biogeographers define biogeography in which our problems lie. How do you define an undefined field?
Blame 19th century malacologist Hermann Jordan. Not much is known about him, other than he was the first person to use the term ‘biogeographie’ (Jordan 1883). Like many of the early users of the term ‘biogeography’, Jordan didn’t define it, meaning that it may have been already in use and its definition known. After all, both plant geography (or phytogeography) and animal geography (zoogeography) were already established fields by the 1880s, and there was no umbrella term to describe them.2 Perhaps the definitions of zoogeography and phytogeography were sufficient enough, but even these terms referred to different practices. A more pragmatic approach is to ask who practised biogeography and work out a definition from there, but there was a snag. Phytogeographers, for example, either studied the distributions of plants (taxonomic groups) or vegetation (plant formations). Zoogeographers referred to either geographical regions or isothermal lines. Which was it? Vegetation such as rainforest consists of many different taxonomic groups. Species within a taxonomic group may be found in multiple vegetation types. Moreover, an isothermal line is based solely on temperature, while a region is based on taxic distributions and geographical barriers. There was no consistency in 1880s biogeography, nor was there a single aim or method. All zoogeographers and phytogeographers wanted to do was look at different aspects of plant and animal distribution. That tradition has remained until the 21st century. Throw in recent discoveries and innovations, such as genetics, deep-sea thermal-vent faunas, phylogenetics and geospatial technology and suddenly the number of aims and methods multiply, but the definition for biogeography remains the same:
The biogeography universe
The multidisciplinary nature of biogeography means it incorporates the aims and methods of various fields (e.g. cladistics, molecular genetics, phenetics, palaeontology and geography). However, much of this book is dedicated to cladistic biogeography, the methods of which are outlined in Parenti and Ebach (2009). Other biogeographic approaches not covered in this book are phylogeography (Avise 2000), palaeobiogeography (Lieberman 2000), ancestral area reconstruction (Ree et al. 2005); human biogeography (Harcourt 2012), island biogeography (MacArthur and Wilson 1967, 2001), ecological biogeography (Ricklefs and Relyea 2013) and conservation biogeography (Ladle and Whittaker 2011). The panbiogeographic approach and its impact on New Zealand biogeography is covered in Chapter 5.
‘The branch of biology that deals with the geographical distribution of plants and animals … the characteristics of an area or organism in this respect’ (OED 2015a).
Surely any branch of biology deals with the geographical distribution of organisms? For example, botany, entomology, ecology, palaeobiology, parasitology, anthropology, bacteriology and mammalogy all deal with a geographical component. So do related fields such as epidemiology, linguistics and human geography. Perhaps seen as a multidisciplinary field, biogeography would make greater sense? After all, what branch of biology doesn’t deal with geographical distribution? Very few indeed.3 In order to understand what biogeography is, we need to see how biogeography is done.
The search for natural biotic areas
Unless you are reading this in outer space, you are currently in – or driving, riding, flying or sailing over – a natural biogeographic region. The region is defined by the organisms that inhabit it – the biota – which have been formed by millions of years of evolutionary, geographical, geological and climatic processes. This includes the barriers that encapsulate the biota, isolating it from other biota. Over time, these biota diversify and become endemic to areas in which they live. If you are standing in a plain near a troop of kangaroos, for example, you are most likely in Australia. But things get moved around. For instance, you may also be standing in a kangaroo enclosure in a zoo. Biogeography is never clear cut; rather it is messy. Animal and plants tend to move around, either naturally or by human-assisted means, such as the introduction of the cane toad in Australia or eucalypts in Spain. Identifying which organisms are part of a natural area is incredibly difficult for several reasons. First, not many people work on more than one taxonomic group. A single taxonomic group does not make a biota. Next is the problem of whether your taxonomic group is actually endemic. It might be introduced or naturally cosmopolitan. The biggest hurdle, however, is determining the relationships between the species and genera under study. Without these, you have no independent source of information about the relationships of your areas. The only way to find out what these relationships are is to find the relationships between the morphological or molecular characteristics of your organisms. For much of the 18th, 19th and 20th centuries, this type of information was unavailable. Not until the 1970s were we able to investigate morphological and genetic relationships using numerical methods. Before then, natural areas were assumed, and never tested, given the lack of an analytical method. Without confirmation of natural areas, much biogeography was descriptive; that is, limited to describing and naming areas. There was, and still is, a dominant component to biogeography – the narrative. Knowing the origins and distributional history of species and their genera has been a mainstay of biogeographers since Linnaeus. Narratives about taxic distribution over time, such as dispersal routes for migrations and extinctions, provide a natural history of the organisms in question. Narration is not descriptive or analytical, nor does it necessarily require a method. Over time, our narratives were synthesised in the conclusions to many taxonomic monographs and phylogenetic studies. Biogeography was nothing more than assuming natural biotic areas and telling stories about the distributional histories of its organisms. That is until 1978, when Donn Eric Rosen, an American ichthyologist at the American Museum of Natural History, made a significant breakthrough in how we find the relationships between biotic areas (Rosen 1978). That breakthrough was a result of a new way of doing biological classification – cladistics – and it completely reformed traditional Linnaean taxonomy.
Cladistics: the search for natural taxa and their relationships
Taxonomy is an often-misunderstood field of research. Unlike ecology, physiology and other biological disciplines, taxonomy has no precise methodology – at least not one that has been translated into an algorithm and may be implemented on a computer. Certainly, books detailing the taxonomic method have been written, with an emphasis on how to classify organisms based on characters, how to name them, and so on (e.g. Blackwelder 1967; Winston 1999). These books, however, are not popular and they are rarely cited or used in teaching taxonomy. At first glance it seems that taxonomy is nothing more than a poor cousin of the other more flamboyant biological sciences – ones that use experimental methods, statistics and super-computers. Taxonomy has also been unfairly labelled an ‘artform’, much to the chagrin of taxonomists. So what is taxonomy and why on Earth would anyone do it?
Taxonomy is the study of classifying organisms into taxonomic groups: namely taxa (e.g. species, genera, families). Catching butterflies in nets, pinning them onto cardboard and giving them a name is what some people think taxonomists (in this case lepidopterists) do all day. Placing specimens into collections, taking them out again and rearranging them into different groups might also be what taxonomists are seen to be doing, but the act of classification is an intricate process that takes place in the mind. Classification is an instinctive process: something we are born with and something that helps us communicate and survive. Without classification we are lost. There would be no day-to-day language with which we can communicate our feelings and facts to others. This is not to say that giving a name to something defines taxonomy. Taxonomy can be done without names. For instance, we may accurately warn others that the barking thing, with four legs, a tail and sharp teeth, definitely bites. The fact that we can see the characteristics of the four-legged biting thing means we are able to identify it from other things that bite, such as the small buzzing black things that live near the swamp. Giving a thing a name is merely a convenience. Being able to identify what that thing is based on the observed characteristics is the genius of thousands of processes that are occurring in our minds. We can even compare the four-legged barking thing with the four-legged purring thing. They share the same characteristics, such as a tail, teeth and four legs, which probably means that they are more like each other than they are to, say, the buzzing biting thing. That is classification – the ability to identify and categorise organisms based on their observed characteristics.
Classification is something that is inherently human; it is part of us, as much as grammar and touch. This is also why most people trained as taxonomists don’t read books on how to do classification, because we have been doing it all our lives. There is no hypothesis to test, nor is there is an experiment. So how do taxonomists know they have got it right? How do they know whether the organisms they classify are in fact natural evolutionary groups? The fact is, they don’t. Taxonomy is not an art and taxonomists are unlike, say, art historians. Good art historians are also adept at classifying artwork and have reliable guides for identifying the authenticity of certain types of oil paintings, which they can classify by artist. This is because art historians have access to provenance – the art catalogues, gallery labels, auction stamps, and so on. If these are missing, they can date the paint or compare brushstrokes with known originals. But taxonomists have no idea if they do in fact have an ‘original’ taxonomic group. Rather taxonomists, through comparison, can tell to a certain degree whether a taxonomic group, or taxon, is based on natural characteristics called homologues, such as feathers in birds and hair in mammals. Taxonomy is classification without provenance, but organisms can be dated (if they are fossils), and we can compare the brushstrokes – namely, the homologues – this is where cladistics comes in.
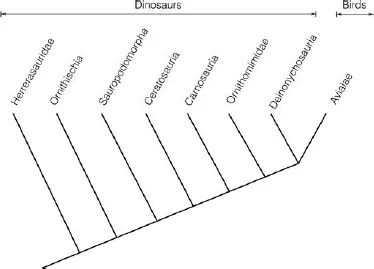
Cladistics is a methodological approach to biological classification in which the relationships among taxa (e.g. species or genera) are determined by shared relationships between homologues. In short, cladistics is a way to find natural taxa. What makes cladistics important is that it takes away the need for dating (in fossils) and relies on patterns of relationships. In this way, cladistics tests whether our classifications are natural and what other taxa they are related to. Cladistics is a tool used in revising potential artificial taxonomic groups. The most famous example is that of the dinosaurs. Originally described from a fossil claw belonging to Iguanodon, dinosaurs were a poorly defined and nebulous collection of taxa. Then in the 1980s, a cladist tested the group and found that some dinosaurs were more closely related to other taxa, such as birds, than they were to anything else, confirming earlier suspicions (Fig. 1.1). The discovery was reported as dinosaurs being the ancestors of birds. The problem with this statement is that dinosaurs are assumed to be a natural taxon. What the classification, or cladogram, is saying in Fig. 1.1, is that birds have a set of homologues that define them as a natural or monophyletic taxon, while the dinosaurs have no diagnostic homologues and are therefore in desperate need of taxonomic revision. One solution would be to place the birds and dinosaurs into a single group, which would make them monophyletic, but it still leaves us with the problem of what diagnostic homologues make members of birds plus dinosaurs a natural taxon. Rather, a taxonomic revision would involve finding shared diagnostic homologues in order to identify natural taxa within Dinosauria. For example, the theropods may be included in the birds, because they share bird homologues. Ornithischians and sauropods may become two separate monophyletic taxa, and so on. In the end, you will have several monophyletic taxa. The name ‘dinosaur’ would simply vanish.
Fig. 1.1. Cladogram depicting phylogenetic relationships among well-known theropods and other dinosaurs. The figure, redrawn from Gauthier (1986, p. 54, Fig. 8), shows that some dinosaurs, such as the theropods, are more closely related to birds (Avialae) than they are to other dinosaurs.
The anatomy of a cladogram
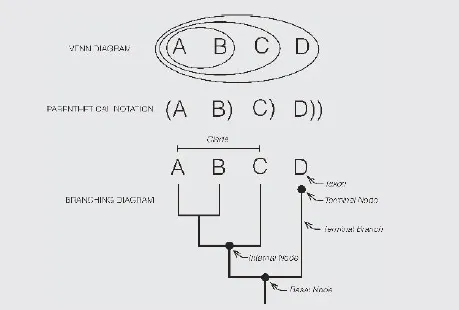
The relationship of homologues is called a homology: namely a relationship. A cladogram is simply a pattern of relationship. The nodes on a cladogram are artefacts of a branching diagram and contain no information. Note that nodes are not present in other types of cladograms, such as in Venn diagrams and in parenthetical notation. A cladogram is evidence for a common history of taxa. Taxa that share a common history are part of a larger natural or monophyletic taxon.
The anatomy of a cladogram.
Removing the dinosaur name would be a correct and practical solution in biological classification, but it would be incredibly unpopular with the public. The term ‘dinosaur’, however, does have a practical use in identifying large lizard-like monsters. A sauropod or a theropod may be identified as a dinosaur. So too can cartoon characters and other non-living and non-fossil objects. Dinosaur, like the terms ‘unicorn’, ‘gryphon’ and ‘leprechaun’, is an identifier and serves a useful purpose within a folk classification. If we return to the question of why birds are a natural group and not dinosaurs, we may ask ‘name the homologues that relate all birds’. You may say ‘beaks’, ‘feathers’ or ‘hollow-bones’. Not all birds fly (think of the emu), so ‘wings’ are not a homologue that relates all birds.4 Now think of a single homologue that relates all members of the Dinosa...