
- 383 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Plant viruses are of considerable interest to the science of biology, and their study has contributed significantly to the elucidation of several mysteries of traditional and molecular biology.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 4
Virus Transport in Plants
C. L. Mandahar and I. D. Garg
Table of Contents
- I. Introduction
- II. Virus Transport
- A. Cell-to-Cell Transport
- 1. General
- 2. Role of Plasmodesmata
- B. Long-Distance Transport
- C. Transport Form of the Virus
- 1. Translocation as Complete Virus Particles
- 2. Translocation as Free Viral Nucleic Acid
- 3. Translocation as Viral Replicative Complex Associated with Cell Membranes
- 4. Translocation as Informosomes
- A. Cell-to-Cell Transport
- III. Role of Viral Genome/Genetic Basis of Transport
- A. Transport Protein
- B. Complementation of Viral Transport Function
- IV. Virus Transport in Xylem
- V. Transport of Viroids
- References
I. Introduction
Viruses enter their hosts through one or more of the following ways: mechanical inoculation through abrasion, and introduction through vectors, dodder, or graft union. Entry is through the epidermis in the case of mechanical inoculation and chiefly by nonpersistent transmission through vectors. Transmission through graft union, dodder, or vectors transmitting the virus in a persistent manner, generally places the virus in the deeper tissue layers of the host. The common sites of virus entry in mechanical inoculation are supposed to be the broken epidermal hairs and/or protoplasmic membranes exposed in the wounded epidermal cells. Abundant plasmodesmata usually occur in cell walls of hairs and between hair cells and the underlying epidermal cells. Virions coming directly into contact with the exposed plasmalemma may enter the cell by pinocytosis. Once inside the cell, virions undergo deproteinization. The released viral nucleic acid then produces one or more polypeptides through translation by using the host protein-synthesis machinery. The viral RNA translation products then take part in replication of viral genomic RNA, which in turn encodes viral coat protein besides one or more species of nonstructural virus proteins.
Upon mechanical inoculation, only a few host cells are infected directly by the virus. Thus, the number of primarily infected cowpea cells upon mechanical inoculation with tobacco mosaic virus (TMV) was estimated to be one in 50,000 to 1,50,000 mesophyll cells.1 For an infection leading to virus disease symptoms, it is essential that virus infection spreads from the initially infected cells to the neighboring cells. This is the cell-to-cell spread of the virus, called slow or short distance transport, and is assumed to take place through plasmodesmata. Systemic infection of a plant can occur only when virus is able to become distributed throughout the plant. This is the long-distance or rapid movement of virus and takes place in phloem. When long-distance transport of a virus is combined with its slow cell-to-cell movement, the two together result in a thorough invasion of the host and lead to rapid appearance of systemic symptoms in infected plants. Thus, cell-to-cell movement of tobacco ringspot virus from inoculated areas to sieve tubes and its long-distance transport in phloem results in systemic infection of soybean plants within 3 to 10 d after inoculation of cotyledons or leaves.2 A third type of virus transport which is intermediate in speed has been suggested by Schneider.3
Virus transport has been reviewed.3-6 Much new and fundamental information has recently become available which has completely transformed our understanding of the various aspects of virus movement in plants.
II. Virus Transport
A. Cell-to-Cell Transport
1. General
Cell-to-cell transport occurs between parenchyma cells and between parenchyma and vascular tissue. Virus particles moving from cell to cell are not the ones originally introduced into the cell(s) by inoculation, but are the new generation of particles produced by multiplication of the original inoculum. The process is presumably repeated for every new cell infected, which explains the slow cell-to-cell movement of a virus. The bulk of the particles seem to remain in the cell in which they are synthesized, while only a limited, but an unknown number, moves on to the adjacent cells. Thus, cucumber mosaic virus (CMV) appeared to be transported from an infected cell to an adjacent cell after considerable accumulation of antigen in infected cells has taken place.7
Virus particles, infectious nucleic acid, and protein coat are nonmobile, and their movement within a cell or to the contiguous cells has obviously been thought to be dependent upon the transport systems operating within the host cell. Cytoplasmic streaming is an important force that brings about the intracellular movement of cytoplasmic entities. Viruses until quite recently were, therefore, also assumed to be translocated passively within the cell, to plasmodesmata, and through plasmodesmata to the adjoining cells with cytoplasmic streaming.3,8
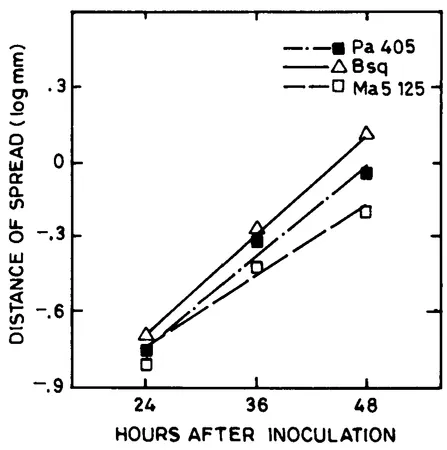
FIGURE 1. Rate of cell-to-cell transport of maize dwarf mosaic virus strain B in mechanicall...
Table of contents
- Cover
- Title
- Copyright
- PREFACE
- THE EDITOR
- CONTRIBUTORS
- ACKNOWLEDGMENTS
- Contents
- SECTION I. THE DISEASED PLANT
- SECTION II. VIRUSES AS PATHOGENS
- SECTION III. DISEASE CONTROL
- INDEX
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Plant Viruses by C.L. Mandahar in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.