
eBook - ePub
Structure and Properties of Cell Membrane Structure and Properties of Cell Membranes
Volume I
- 238 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Structure and Properties of Cell Membrane Structure and Properties of Cell Membranes
Volume I
About this book
This book provides in-depth presentations in membrane biology by specialists of international repute. The volumes examine world literature on recent advances in understanding the molecular struc-ture and properties of membranes, the role they play in cellular physiology and cell-cell interactions, and the alterations leading to abnormal cells. Illustrations, tables, and useful appendices com-plement the text. Those professionals actively working in the field of cell membrane investigations as well as biologists, biochemists, biophysicists, physicians, and academicians, will find this work beneficial.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
BiowissenschaftenSubtopic
BiologieChapter 1
THE EVOLUTION OF MEMBRANE MODELS
TABLE OF CONTENTS
I. | The Beginning of an Evolution |
II. | The First Branch |
III. | The Second Branch |
IV. | The Third Branch |
V. | The Sucker |
VI. | A Vision |
References | |
I. THE BEGINNING OF AN EVOLUTION
Before electron microscopy made it possible to observe membranes, it was theoretically assumed that membranes are present at the surfaces of the cells and the cell nucleus to account for the difference in the composition of the media on the two sides of these boundaries. The membranes were conceived of as forming barriers that controlled the exchange between these media, passively due to the physical properties of the membranes.
The observation by Overton1 in 1899 that substances that are soluble in nonpolar solvents permeated the surface of plant protoplasts faster than water-soluble substances led him to propose that the surface of cells is impregnated by a substance that might be similar to a fatty oil.
This observation drew the attention to the lipids as possible basic components of the cell surface, and it led to the first membrane model proposed by Gorter and Grendel2 in 1925, according to which a single lipid bilayer covers the cell surface (Figure 1).
The observation by Danielli and Harvey3 in 1935 that the surface tension at the cell surface is too low to correspond to that of a lipid surface led to the membrane model of Danielli and Davson4 in which a layer of protein molecules was added to each side of the lipid bilayer. The proteins would act as surfactants reducing the surface tension. In this second membrane model, the lipid bilayer is thus sandwiched between two protein layers that were originally assumed to consist of globular proteins, while later the polypeptide chains were considered to be unfolded (Figure 2).
The role of the proteins was to stabilize the lipid bilayer, and they were not considered to play any specific role by contributing to the permeability properties of the membrane. Addition of possible charge interactions between the proteins and the lipids made the model appear somewhat more sophisticated.
The further evolution of membrane models reminds one in certain aspects of the evolution of year models of automobiles. Like the automobile manufacturers who have realized that it is commercially expedient to change the shapes of automobiles only gradually, because too drastic changes have led to catastrophic declines in sales, we find that only gradual changes in the membrane models have been generally accepted, while extensive modifications have been ignored.
Electron microscopic analysis of the structure of cells led to the discovery of membranes as existing as discrete structural entities. It was found that membranes are not confined to the boundaries of cells and of the cell nucleus, but that they constitute the dominant structural components of practically all cell organelles in the cytoplasm. The cytoplasmic membranes were thus discovered starting with the membranes of the outer segment disks in photoreceptors by Sjöstrand,5,6 followed by the discovery of membranes in the chloroplasts by Steinmann,7 in the mitochondria by Palade8 and by Sjöstrand,9 in the Golgi apparatus by Dalton and Felix10 and by Sjöstrand and Hanzon,11 and in the basophilic cytoplasm, the ergastoplasm, that was found to correspond to the endoplasmic reticulum observed in whole cells in tissue cultures by Porter and Kallman.12
The cytoplasmic membranes were proposed by Sjöstrand9 in 1953 to play a more competent role functionally than to form a barrier. This point of view was expressed by proposing that “the intracellular cytoplasmic membranes may represent a fundamental principle of organization of the cytoplasm.”
However, the generally accepted concept was that membranes function as barriers, and the function of the cytoplasmic membranes including the mitochondrial membranes is to delimit compartments. The interest was, consequently, focused on the lipid bilayer structure of membranes.
For many years, the triple-layered pattern observed in electron micrographs of crosssections of membranes was accepted as confirming the Danielli-Davson membrane model, and the universal presence of this pattern led to the concept that all membranes are structurally identical. For as many years, the proteins that constitute the major part of the mass of most membranes were ignored except those that contributed to the sandwich structure. They could easily be ignored because they were lost during the preparation of the specimens, although this was not acknowledged.
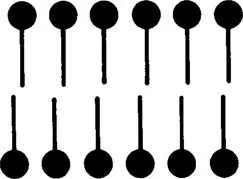
FIGURE 1. The lipid bilayer model for the plasma membrane proposed by Gorter and Grendel.2
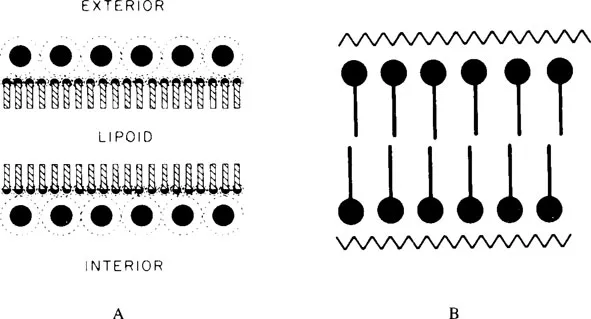
FIGURE 2. (A) The original model for the plasma membrane proposed by Danielli and Davson;3 (B) the modified model published in 1952 by Davson and Danielli. (From Davson. H. A. and Danielli, J. F., The Permeability of Natural Membranes, Cambridge University Press, London, 1952. With permission.)
With the advent of high-resolution electron microscopic analysis of the structure of cells, membrane models could be based on direct observations. However, not all membrane models that were proposed from now on were based on such observations. Some were, instead, deduced on theoretical ground. Of those that were based on observations, it is possible to distinguish those in which the observations have been interpreted objectively without any preconceived concept regarding the structure of membranes and those in which the interpretation has been based on a dogmatically assumed basic structure of membranes that applies universally to all membranes.
We can, therefore, distinguish between different approaches that we will represent by different branches of our evolutionary tree. This tree also has a sucker.
II. THE FIRST BRANCH
The first branch is contributed by membrane models that have been developed on the basis of an objective evaluation of observations. On this branch, we find a model that represents a first attempt to incorporate proteins in the structure that were not associated with stabilizing the sandwich structure. The plasma membrane was found to be asymmetric with a thicker stained layer located at the cytoplasmic surface. The thickness of this layer could be accounted for by an accumulation of proteins at this surface.13 As shown in Figure 3, this layer was assumed to consist of globular protein molecules.
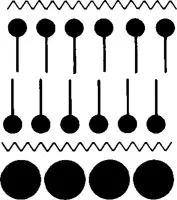
FIGURE 3. A layer of globular protein molecules was added by Sjöstrand13 in 1960 to the cytoplasmic surface of the Danielli-Davson model to account for electron microscopic observations.
During the 1960s and 1970s, the proposed membrane models reflect more concern about the role the proteins may play in the structure of membranes. It eventually became obvious to some researchers that the technique to prepare tissues for electron microscopic analysis that had been so useful in making it possible to discover the cytoplasmic membranes was not satisfactory for an analysis of the membrane structure at a molecular level, because the preparatory technique led to an extensive denaturation of the proteins.
It had thus become obvious that the triple-layered structure of membranes as revealed in electron micrographs could be caused by a denaturation of the membrane proteins, because this pattern appeared irrespective of the lipid content of membranes and, in the case of mitochondrial membranes, even after extraction of practically all lipids.14 This pattern could, therefore, not be used as a criterion for the existence of a continuous lipid bilayer if it was not known that such a bilayer existed.
Attempts were now made to improve the preparatory techniques with the aim of limiting sufficiently the extent to which the proteins were denatured to make the protein molecules retain their globular conformation. Several preparatory methods were developed and refined during the 1960s and 1970s that made it possible to collect new information regarding the structure of membranes.
An intermediate step in this development involved a slight modification of the conventional technique that, however, led to pictures of membranes showing a particulate structure instead of a triple-layered pattern. Cytoplasmic membranes seemed to consist of a layer of globules that were first interpreted to consist of lipids but later were considered to be globular proteins.15,16 A membrane model was proposed according to which globular protein molecules were embedded in a lipid bilayer in such a way that they extended across the bilayer and were exposed at both surfaces of the membrane (Figure 4).
This observation, which later could be shown to involve a fixation artifact, was encouraging because it seemed to reveal how a globular protein molecule would appear in an electron microscopic picture, and it showed that a sufficiently high resolution could be achieved on thin sections to make it possible to observe individual protein molecules when in their globular conformation. ...
Table of contents
- Cover
- Title Page
- Copyright Page
- Table of Contents
- Chapter 1 The Evolution of Membrane Models
- Chapter 2 Protein-Protein Interactions in Cell Membranes
- Chapter 3 Lateral Mobility of Proteins in Membranes
- Chapter 4 Lateral Diffusion of Lipids
- Chapter 5 Topological Asymmetry and Flip-Flop of Phospholipids in Biological Membranes
- Chapter 6 Membrane Fluidity: Molecular Basis and Physiological Significance
- Chapter 7 Lipid Dependence of Membrane Enzymes
- Chapter 8 Protein-Lipid Interactions in Biological Membranes
- Chapter 9 Basement Membrane Structure, Function, and Alteration in Disease
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Structure and Properties of Cell Membrane Structure and Properties of Cell Membranes by Gheorghe Benga,Benga in PDF and/or ePUB format, as well as other popular books in Biowissenschaften & Biologie. We have over 1.5 million books available in our catalogue for you to explore.