![]()
CHAPTER 1
Cilia in Brain Development and Disease
Gilbert Lauter, Peter Swoboda* and Isabel Tapia-Páez
INTRODUCTION
Brain Development
The notion that the human brain is the most complex object in the universe may be a narcissistic statement. However, the roughly 1.3 kg of human brain does grant its bearer rather astonishing capabilities, ranging from the subconscious control of bodily functions and complex motor skills to consciousness, emotions, memory, language and reasoning. Still, the complex structure of the adult human brain develops gradually from a simple field of neural cells. Insights into embryonic neural development might thus contribute to a better understanding of how neuronal interaction can result in all the executive and cognitive functions carried out by the human brain. Vertebrate brain development follows an intrinsically orchestrated sequence of events that are highly conserved between species. During neurulation, the ectoderm-derived planar neural plate bends, folds and finally closes to form the neural tube. Morphologically distinguishable brain vesicles start to appear as the neural tube becomes progressively subdivided along its rostro-caudal axis. The well-known three-vesicle stage with the prominent bulges of the fore-(prosencephalon), mid-(mesencephalon) and hindbrain (rhombencephalon) thus forms. Subsequently it is replaced by the five-vesicle stage, which is derived through further subdivision of the pros- and rhombencephalon. Interestingly, this sequence of neuromeric bulges occurs in all vertebrates at a certain developmental stage and results from zones of high mitotic activity located at the center of each vesicle (Bergquist, 1952; Bergquist and Källén, 1954; Källén, 1952). Taking into account the actual curvature of the developing brain, Bergquist and Källén demonstrated that mitotic zones are arranged transversely. This transverse arrangement of so-called neuromeres along the brain axis argues in favor of a segmental organization of the brain, thereby questioning the back then predominating columnar model, which stated that more or less the entire brain is organized into longitudinal, functional columns similar to those found in the spinal cord (Herrick, 1910). During subsequent development, transverse and later-forming longitudinal proliferation zones intersect to partition the brain into a patchwork of mitotic areas or so-called migration areas, where each area constitutes the future site for differentiation and migration (Bergquist and Källén, 1954). Surprisingly, the topography of migration areas is highly conserved between species and therefore suitable for establishing homology relationships on the basis of shared origin instead of similar function, a concept summarized by the term field homology (Nieuwenhuys, 2009; Puelles and Medina, 2002). The conserved patchwork of migration areas can be seen as an example of a common vertebrate bauplan (“construction plan”) for brain development. With the advent of modern molecular techniques, even more support for segmental brain organization and insights into the framework behind the common vertebrate bauplan, were gained.
During development, regional specification of the brain arises progressively through repeated subdivisions of larger areas into smaller ones. The underlying patterning mechanisms are similar to those employed for the development of other body parts. In general, transient cues of positional information are conveyed via signaling molecules and instruct small cell populations to act as self-organizing centers. In turn, these so-called local organizers or signaling centers control the release of morphogens to which the surrounding tissues respond. The initial, transient positional information is tightly coupled to early events in axis formation during which morphogen gradients of Fibroblast Growth Factor (FGF), WNT and retinoic acid are set up across the anterior-posterior aspect of the developing brain. Subsequently, the dose-dependent response of the receiving tissue results in the expression of transcription factors (TFs) that mark broad regional identities (Sanes et al., 2006). Organizers typically arise at the boundary between regional markers (Macdonald et al., 1994). One prominent example of an early organizer is the mid- to hindbrain boundary (MHB), located at the interphase between Orthodenticle Homeobox 2 (OTX2) and Gastrulation Brain Homeobox 2 (GBX2) expression. The MHB arises through mutual repression between OTX2 and GBX2, which eventually leads to a sharp boundary and maintained expression of FGF8. In turn, positional information is mediated via the secretion of FGF8 to adjacent areas of the future mid- and hindbrain (reviewed in Raible and Brand, 2004). Thereby surrounding cells acquire different cell fates, depending on the concentration of a received morphogen and on their competence. Similar organizing centers, albeit with different molecular players at work, are set up at various positions along the neuraxis (reviewed in Cavodeassi and Houart, 2012). For example, the zona limitans intrathalamica (ZLI) is marked by the prominent expression of Sonic Hedghog (SHH) between the future rostral and caudal thalamus (reviewed in Scholpp and Lumsden, 2010). The anterior neural ridge found at the anterior-most end of the developing brain, is essential for induction and development of the telencephalon via regulation of the WNT signaling pathway (Houart et al., 1998; Shimamura and Rubenstein, 1997). By contrast to these centers controlling cell fate specification along the anterior-posterior axis, the floor plate is a prime example for a local organizer acting primarily along the dorsalventral axis. This part of the ventral midline constitutes a continuous source for secreted SHH, which eventually leads to the establishment of different neural progenitor zones by differential SHH responsiveness (Briscoe et al., 2000; reviewed in Caspary and Anderson, 2003).
In summary, pan-neural patterning events of axis determination set up the major brain areas by broad expression of regional TFs. Subsequently, the concerted action of organizing centers subdivide larger regional identities into smaller histogenetic fields marked by the expression of a unique transcription factor specification code. In vertebrates, these specification codes have been largely conserved across species and constitute the genetic framework behind a common vertebrate bauplan for brain development (Ferran et al., 2007; Hauptmann and Gerster, 2000; Lauter et al., 2013; Puelles and Rubenstein, 2003). Eventually, progenitor domains form and secondary histogenetic processes like proliferation, migration, differentiation, synaptogenesis and apoptosis follow an exact spatio-temporal sequence to shape the complex structure of the brain with its various specialized brain functions.
Brain Anatomy
Vertebrates inhabit very diverse environmental niches. It is therefore not surprising that their brains have evolved various specializations. Still, mature brains are the result of a sequence of highly conserved developmental steps and therefore, they share major brain subdivisions (see above). Specializations can thus be seen as deviations from a common, albeit hypothetical, archetype by further evolution and elaboration of only certain parts. For example, a dorsal pallium can be found in all vertebrates but in primates it tends to enlarge substantially, culminating in the human cortex (Striedter, 2005). Similarly, a cerebellum can be distinguished in all vertebrates, but the cerebellum of animals that move in a three-dimensional space, like birds and fish, often displays greater complexity. A simplified archetype proposes shared forebrain regions of the telencephalon, like the pallium and subpallium in addition to the olfactory bulbs and the hypothalamus. The diencephalon of the forebrain is built by the epithalamus together with rostral and caudal thalami, followed by pretectal areas. The midbrain consists of the tectum and tegmentum, whereas the cerebellum, pons and medulla represent the major parts of the hindbrain. It should be noted, however, that such a simplified archetype is of a hypothetical nature and it represents a principle that integrates conserved, universal features. It primarily functions as a template to facilitate cross-species comparisons (Striedter, 2005).
Inevitably, these universalities extend beyond the regional morphological structures and they can also be found at the cellular and molecular level. All brains are constructed with the same cellular building blocks, derivatives of neurons and glia together with a vascular supply. Furthermore, cells group together and they form defined aggregates of similar organization as seen in laminae, brain nuclei or loose, reticular formations. At the subcellular level, a key universal feature of the brain is the presence of specialized contact points for communication, referred to as the synapses. At the synaptic cleft the propagation of electrochemical signals is regulated in a way that allows neurons to transmit, receive and integrate information. The computational power of the synapses relies on the exact spatial and temporal organization of molecules like scaffolding proteins, neurotransmitters and neuromodulators. This molecular machinery is highly conserved. The same set of neurotransmitters enables neurons to perform their typical functions not only in all vertebrates, but also in invertebrates. Despite the wealth and detail of knowledge of many of the components of the brain, some uncertainty prevails about how the coordinated (inter-)action of all its parts guarantees brain function.
Brain Physiology
From a reductionist point of view, the brain represents a central hub, which coordinates interactions of the individual with the environment to ensure survival-promoting behavior. In order to decide upon an adequate behavioral response, information about the state of the environment has to be gathered.
Sensory perception of the environment starts when a specialized sensory cell picks up an adequate stimulus via the corresponding sensory molecule or receptor. During subsequent steps this signal is turned into an electrochemical (neuronal) signal, which is propagated to the respective brain areas. Interestingly, receptors are often concentrated and exposed to the environment in specialized subcellular structures that rely on cilia (see below). In most vertebrates the sensory system commonly includes the perception of light, sound, taste, odor, touch, heat, posture and pain. Further, depending on the environment and the concomitant selective pressure, animals have evolved astonishing specializations resulting in extreme sensitivity to the adequate stimulus, sometimes even reaching physical limits (Frings, 2012). Following receptor activation, a complex interplay between second messenger and ion channel dependent mechanisms results in signal amplification and the production of electrical charges. The generation of action potentials follows an all or nothing principle in the sense that sensory cells will either not respond at all or generate full-fledged potentials subsequent to stimulation. From any given stimulus the sensory system tries to acquire information not only about the type of stimulus received, but also about its duration, intensity and localization. The wealth of information sensed simultaneously by all the senses is truly enormous, keeping in mind that furthermore, the brain is constantly fed with information about the current body status through other systems. In addition, information can be compared to past, memorized events and the consequences of future actions can be assessed. But even though various information sources are of totally different nature, the readable output transmitted in the brain almost always consists of propagating action potentials, tiny localized bursts of electrical charges. The uniformity of action potentials implicates that information must be encoded by the timing and frequency of discharges. Frequency modulation seems to be the general modus operandi for encoding information in the brain (Frings, 2012). Albeit the format of exchangeable information is uniform and while we have good knowledge about the basic functional units, we still lack a fundamental understanding of the steps leading from cellular chemistry to cognition.
However, some typical features are associated with the appearance of cognitive functions. Less evolved brains tend to have fewer areas connected between primary sensory and executive structures. With the evolution of higher cognitive functions, more and more cortical areas are added into pathways. Thus, the addition of cortical areas that are highly inter-connected but only indirectly linked to sensory and executive entities seems to be essential. At the same time the connectivity of participating areas does not follow a linear, hierarchal logic. Instead of a single command center where everything converges, information seems to be processed in parallel and reciprocally in different brain areas by varying cell assemblies (Singer, 2012). It seems that convergence in time, not space, is essential for letting us enjoy such amazing brain functions as for example reading.
Cilia in the Brain
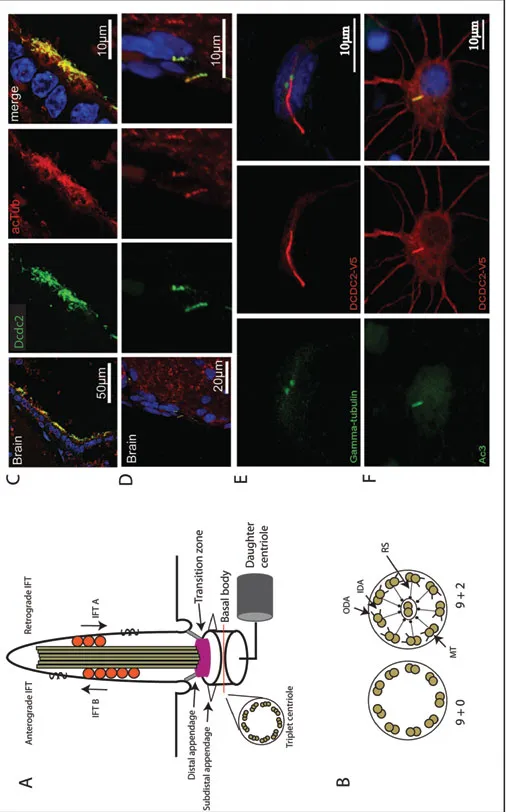
Cilia are hair-like cell protrusions found on many different cell types within the eukaryotic kingdom (Piasecki et al., 2010). The cilium consists of a microtubule based core structure, the axoneme, surrounded by a specialized ciliary membrane (Figure 1A). The axoneme originates from the basal body positioned close to the cell membrane as a ring of nine microtubular doublets (Figure 1B). Primary and sensory cilia typically display this 9 + 0 configuration, whereas motile cilia contain an extra pair of microtubules in the middle of the ring and therefore adopt a 9 + 2 configuration (Figure 1B). Auxiliary structures like outer and inner dynein arms together with radial spokes are required to ensure motility. Active, motor-protein driven intraflagellar transport (IFT) is essential for the formation and maintenance as well as for the function of the ciliary compartment. Despite their wide distribution and functional importance, cilia have long been overlooked. This cellular organelle is present on most polarized cell types in the human body and is especially abundant in the brain (Sarkisian and Guadiana, 2015), where motile cilia exist on the ependymal lining of the ventricular walls (Spassky, 2013) and primary cilia are present on most neuronal cell types ranging from progenitor cells to differentiated neurons and astrocytes (Arellano et al., 2012; Bishop et al., 2007; Cohen, 1987; Dubreuil et al., 2007; Fawcett, 1954; Fuchs and Schwark, 2004; Tong et al., 2014). Examples for functional contributions of cilia can be found for most, if not all, morphogenetic processes active during brain development (Guo et al., 2015). The high number of brain conditions associated with human ciliopathies underscores the importance of proper cilia function for normal brain development.
Figure 1. Cilia in the brain. (A) Schematic drawing of the structure of cilia showing the daughter centriole, the basal body, the transition zone, the axoneme with anterograde and retrograde intraflagellar transport (IFT) and a cross section of the basal body depicting the nine triplets of microtubules. (B) Cross section of the ciliary axoneme showing the structure of 9 + 0 primary cilia and 9 + 2 motile cilia, respectively. Outer dynein arms (ODA), inner dynein arms (IDA), microtubule doublets (MT) and radial spokes (RS) are depicted. (C) and (D) The ciliary marker acetylated tubulin (acTub) and the dyslexia candidate protein Dcdc2 stain the ciliary axoneme in mouse ependymal cells of the third brain ventricle (C) and cranial pia mater (D). Original pictures were kindly provided by Andrea Bieder and reproduced with permission (from Schuele...