1
Pregnancy Loss and Fecundability in Women
Darryl J. Holman and James W. Wood
Compared to other mammals, reproduction in women is characterized by low lifetime fertility, a slow pace of reproduction, and a large investment in each offspring. The reproductive span begins relatively late in life; births tend to be spaced at increasingly longer intervals until they stop altogether. Many women then live well beyond ages at which reproduction is possible, something that is rarely seen in other mammals. Making sense out of this pattern of reproduction has been a goal of anthropologists, demographers, physiologists, and evolutionary biologists.
In this chapter, we examine one aspect of this pattern: the way that births are spaced across the female reproductive life span. The timing of births is a complex outcome of many physiological, cultural, and behavioral factors, but our focus will be on two of the components that play an important role in shaping female fecundity and birth spacing. These components are pregnancy loss, which is defined as the loss of any product of conception prior to birth, and fecundability, defined as the monthly or cycle-wise probability of conception. We pay particular attention to some of the methodological difficulties that are encountered in trying to measure fecundability and pregnancy loss, and how these difficulties have limited or distorted our understanding of age-related changes in female fecundity. We propose new methods to overcome the difficulties and apply the methods to data collected in rural Bangladesh.
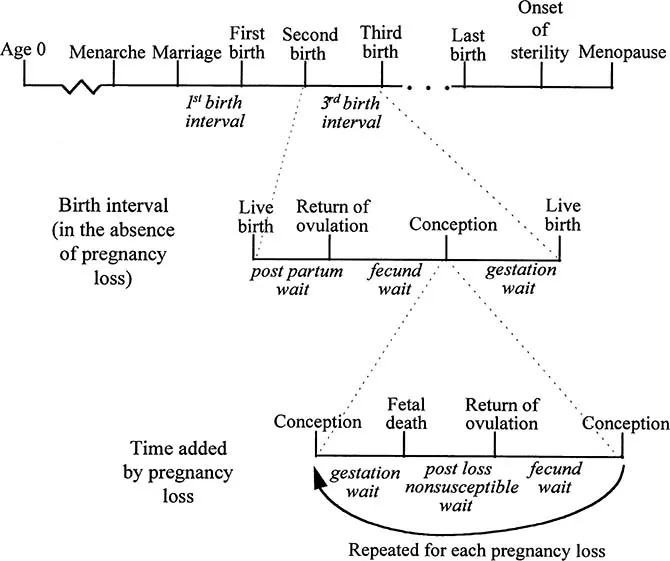
The physiological changes associated with menarche and menopause explain much of the lower levels of natural fertility at the extremes of the reproductive span. Between these two points substantial changes in fertility can be observed as an increase in the average length of birth intervals. A universal finding from studies among natural fertility populations is that female fecundity initially increases to a peak in the early twenties and then declines with a woman’s age.1 These age-related changes in birth spacing can be fruitfully explored by dividing the reproductive life course into a series of smaller components (Figure 1.1). At the top level, the reproductive life course is a series of events, including menarche, menopause, and a number of birth intervals. The time from marriage to first birth defines the first birth interval, and each subsequent birth defines the start of a new birth interval. Each birth interval can be subdivided into four meaningful events separated by three waiting times (Figure 1.1, row 2). Alive birth is followed by a waiting time until the return of fecundity. This is followed by a. fecund waiting time to the next conception, which is some number of months or menstrual cycles until a conception occurs. Finally, a conception is followed by a period of gestation which, in the absence of pregnancy loss, terminates in a live birth.
The role that pregnancy loss plays in shaping birth intervals can be understood from the third row of Figure 1.1. At the time a pregnancy is lost, some period of gestation leading up to that point has been added to the current birth interval. The distribution of these partial gestations is determined by the gestational age-specific risk (or hazard) of pregnancy loss.
Figure 1.1 A schematic of events that occur in the female reproductive life course. The vertical lines represent events, and events are separated by waiting times (after Bongaarts and Potter 1983).
After a pregnancy is lost, three new waiting times are added to the birth interval. The first is a post loss nonsusceptible wait, which is a period of time in which a woman is not susceptible to conception. A pregnancy lost immediately after conception will add little to this waiting time. Wilcox and colleagues (1988) report the lengths of menstrual cycles following 43 subclinical pregnancy losses that were detected, on average, by day 11. Menstrual cycles were lengthened by an average of two days, which represents the combined effects of the length of gestation that preceded the loss and any delays added through the follicular phase of the following cycle. One third of the subjects who experienced an early pregnancy loss conceived in the next cycle (compared with 25% for the study on the whole), so it appears that these early losses do not lead to a high probability of anovulation in the following cycles. Studies of resumption of menses in women who never breast-feed provide information about the other extreme. About six weeks lapse from a live birth to first ovulation in non-breastfeeding women (Gray et al. 1987; Jones 1989). Aside from these two extremes, little is known about the distribution of times from pregnancy loss until the return of fecundity.
The second time added by a pregnancy loss is a new fecund waiting time until the next conception. There has been little investigation of this waiting time, but it is reasonable to suppose that it is similar to the initial waiting time to conception in the second panel. Finally, a new period of gestation follows the fecund waiting time.
Within one birth interval, a woman may become pregnant and lose the pregnancy any number of times. The total number of pregnancies lost within a single birth interval will depend on the probability of loss for each conception. If this probability is high, multiple pregnancy losses can occur within one birth interval, each time adding three new waiting times and resulting in a substantially lengthened birth interval. On the other hand, if the probability of pregnancy loss is very low, then the additional waiting times are rarely added even once per birth interval. Wood (1994:261) gives a mathematical treatment of the relationship between pregnancy loss, the waiting times added by pregnancy loss, and the distribution of birth interval lengths.
Throughout the rest of this chapter, we examine in more detail three of the components that make up birth intervals. First we examine fecund-ability, which determines the fecund waiting time to the next conception. Then we examine the distribution of gestational ages at which pregnancies are lost and the overall probability of pregnancy loss. As will be clear from the discussion that follows, all three components must be treated together for a complete understanding of any one component.
Fecundability
Fecundability is defined as the monthly or cycle-wise probability of conception for a couple that is sexually active, not contracepting, and capable of getting pregnant (Gini 1924). This probability directly determines the waiting time to next conception. In the simplest case, if fecundability is homogenous among and within women, then it is simply the inverse of the mean waiting time to conception (Sheps and Menken 1973).
A number of different methods have been devised for estimating fecundability It has been estimated from the fraction of couples conceiving in a month (Gini 1924; Henry 1972; Potter 1961; Sheps 1965; Tietze et al. 1950), waiting times to a recognized conception (Henry 1972; Jain 1969; Majumdar and Sheps 1970; Potter and Parker 1964; Sheps 1964; Sheps and Menken 1973; Strassmann and Warner 1998), and waiting times from marriage to the first birth (Bongaarts 1975; Gini 1924; Wood et al. 1994). At the most detailed level, fecundability is found as a function of daily probabilities of conception given coitus for each day near ovulation (e.g., Barrett and Marshall 1969; Weinberg et al. 1994).
A fundamental difficulty with all measures of fecundability is that early pregnancies cannot be detected by any current noninvasive technology. Thus any pregnancies that terminate before being detected lead to underestimation of fecundability. Demographers have defined fecundability in a number of ways in order to take into account this difficulty. Total fecundability is defined as the true monthly or cycle-wise probability of conception, apparent fecundability is the monthly or cycle-wise probability of conception using a particular technology to detect a pregnancy, and effective fecundability is the monthly or cycle-wise probability of conception that results in a live birth.
Total fecundability cannot be measured by existing technology; yet, it is conceptually the most important measure because it completely disentangles fecundability from the effects of pregnancy loss (which is a measure of mortality). We will revisit issues of how total fecundability and pregnancy loss are confounded as well as age-specific total fecundability after we examine pregnancy loss in more detail. Both apparent and effective fecundability are measures that confound total fecundability and pregnancy loss. Effective fecundability discounts pregnancy loss altogether. Apparent fecundability is more difficult to interpret, since we must consider the technology being used for detecting pregnancies and how early pregnancies are detected by the method.
The effect of age on apparent fecundability is fundamental for an understanding of birth spacing. The observed pattern of age-specific apparent fecundability based on earlier data shows a rapid increase from zero in the teens to a peak in the early twenties. Thereafter fecundability declines steadily with age until reaching zero in the mid-forties (Wood 1994). It is important to keep in mind that the age-specific decline in apparent fecundability may be biased by an age-related increase in early (i.e., undetectable) pregnancy loss. The extent of this bias can only be known by measuring total fecundability.
Pregnancy Loss
In trying to evaluate the effect of pregnancy loss on the human life course, we run into the same difficulty that we had for fecundability. Since some pregnancies may terminate before they can be detected, empirical studies underenumerate the true number of pregnancies and therefore the number of pregnancies that are lost. Again, the extent of this underenumeration depends on the characteristics (particularly the sensitivity) of the methods used to diagnose pregnancy. For this reason, it will prove useful to use the modifiers total and apparent with pregnancy loss in the same sense they were used for fecundability. Total pregnancy loss refers to all pregnancy loss from conception through term, and apparent pregnancy loss refers to losses that occur after a pregnancy is diagnosed.
As with fecundability, comparisons among studies of pregnancy loss are problematic. Even so, it is helpful to examine the results of broadly similar studies, and to see the effects that the study methods have on apparent pregnancy loss.
Community Surveys
The earliest large-scale population-based studies of pregnancy loss were community surveys that relied on subjects’ self-reports of pregnancy. The apparent probability of pregnancy loss found in these studies are in the range of 12% to 15% per conception. Details and summaries of some of these studies can be found elsewhere (Boklage 1990; Kline et al. 1989; Leridon 1977; Wood 1994). Determining pregnancy by self-reports is not a very sensitive assay method, so that all of these community-based studies grossly underestimated the probability of total pregnancy loss. Still, when properly analyzed, these studies provide a useful picture for the risk of pregnancy loss at later gestational ages.
hCG-based Studies
A number of biochemical changes that take place early in pregnancy can be used to diagnose pregnancy (reviewed in Grudzinskas and Nysenbaum 1985). To date, the only method that has been refined, validated, and used extensively for studies of pregnancy loss involves assays for the hormone human chorionic gonadotropin (hCG) in maternal blood or urine. These hCG-based methods show a high sensitivity and are capable of detecting pregnancies before the end of the second week after fertilization. Even so, the most sensitive assays of this type cannot detect pregnancies until about seven days after conception (Lenton 1988) and fail to detect half of all true pregnancies up to about 10–14 days after ovulation (Holman et al. 1998).2
Another important characteristic of a pregnancy assay is its specificity, defined as the probability that the assay will correctly diagnose a nonpregnant woman as not pregnant.3 Extremely sensitive pregnancy assays tend to have lower specificity because they pick up low levels of background hCG that occur naturally in non-pregnant women (Alfthan et al. 1987; Armstrong et al. 1984; Stenman et al. 1987). Some hCG-based assays have low specificity for another reason: they cross-react with molecules that are similar to hCG, particularly luteinizing hormone (LH). High specificity is extremely important for studies of pregnancy loss, as many assays will be carried out for every early pregnancy loss detected, so that even a small false-positive rate will bias upward the probability of pregnancy loss (Weinberg et al. 1992).
The first large-scale study of apparent pregnancy loss using hCG-based pregnancy assays was that of Miller and colleagues (1980), who measured urinary hCG concentrations in first morning urines taken every other day over the luteal phase of 197 women (mean age 27.5 years) who were discontinuing nonhormonal contraception. The hCG assay they used showed “very little” cross-reaction to LH. Unfortunately, additional details on specificity were not provided, and they did not use a pool of non-pregnant women to test the specificity of the assay under the conditions of study. The limit of detection of the assay was 10 IU/L hCG, and their criterion for pregnancy was a concentration above 20 IU/L hCG in two successive samples or a single sample over 50 IU/L hCG. The probability of pregnancy...