
- 186 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Developments in Lymphoid Cell Biology
About this book
In this volume the author by no means attempted to give a comprehensive view of the myriad of recent developments in immunology. The author has attempted, after introducing sufficient background in chapter 1, to highlight certain areas of lymphoid cell biology which are given less attention in current thinking but which seem to the author to offer exciting prospects for research. In other cases, the important topics of transplantation and tolerance are looked at from points of view which are somewhat less conventional in the hope that these aspects may stimulate others to look at the yet unsolved problems in these areas. In making the selection of topics and indeed in the selection of references within each chapter, the contributors and the author have attempted to select those which offered what seemed to be the best sources of information. They do not wish to minimize the important contributions of others but are guided by constraints of brevity and cogent expression.
Tools to learn more effectively

Saving Books

Keyword Search

Annotating Text

Listen to it instead
Information
Chapter 1
INTERACTIONS BETWEEN LYMPHOID CELLS
A. Arthur Gottlieb
TABLE OF CONTENTS
Introduction
Origin of Lymphoid Cells
Clonal Selection
Antigen-binding cells
Nature of Lymphocyte Receptors
“T” Cell Receptor
Isoantigens on “T” Cells
Is There a Marker Distinctive for “B” Cells?
“T” Cell Subpopulations
Thymosin
“T” Cell-“B” Cell Interaction
The Carrier Effect and the Role of the Carrier in the 2 Immune Response
Suppressive Effects of “T” Cells
“T” and “B” Cells in the Human Lymphoid System
References
Introduction
The ability of an organism to respond to foreign substances in its environment represents a basic biological requirement for survival. In higher forms of life, elaborate mechanisms have evolved permitting defense against foreign substances in many subtle ways. The immune system represents a major way in which materials foreign to one’s own body components are recognized. The major responsibility for the execution of this form of adaptive response goes to the lymphoid system.
The complexity of the immune system is manifested at many levels. In the first place, immunity can be mediated by cells which have the specific capability of recognizing a given antigen, or alternatively by the production by other highly differentiated cells of a specific antibody. Moreover, there is heterogeneity in both kinds of response — not all cells which are sensitized are capable of the same degree of responsiveness. The antibody found in serum is heterogeneous with respect to its ability to bind antigen. Furthermore, antibody may bind tightly to the surfaces of certain cells conferring on that cell the ability to recognize antigens for which the cell lacks the definitive capability of response itself. Finally, there are interactions between different cells through cell contact or through the generation of specific substances which are signals for certain cells to respond in certain ways.
Origin of Lymphoid Cells
A basic outline of the cells involved in the various activities of the immune system is given in Figure 1.
All cells of the lymphoid system have their origin in reticulum cells (stem cells) which are predominantly found in the bone marrow, though they also exist in other lymphoid tissues.
The reticulum cell can give rise to a large variety of cells and is therefore properly regarded as pluripotent. Cells leading to or mediating immune mechanisms are nearly all members of the lymphocytic series of stem cell progeny. The exception is the tissue monocyte, or macrophage, which is generally not thought to arise from or to give rise to any of the other immunocompetent cells. Monocytes apparently arise directly from the reticulum cell.
The group of cells (referred to as immunocompetent) which are either capable of reacting with antigen or which actually mediate immune responses do so owing to acquisition, during differentiation from their stem cell ancestors, of mechanisms which can recognize the specificity of a given antigen. Such mechanisms are to be distinguished from the nonspecific responses of phagocytic cells such as macrophages to foreign substances. One of the key questions in the evolution of the immune response is: At what point in evolution does a purely phagocytic mechanism evolve into unique cell-mediated responses against specific antigens?
Stem cells originate in the embryonic yolk sac and migrate to hematopoietic colonies in fetal liver and bone marrow, then onto other lymphoid tissues where further differentiation occurs.1, 2 and 3 The commitment to lymphoid or myeloid differentiation (as in the spleen) apparently is regulated by inductive factors present in the microenvironment of these organs.4
Within the lymphoid system, two routes are possible. These are best seen in the bird, where there are two distinct primary lymphoid organs —
the thymus and the bursa of Fabricius. Removal of the bursa from a newborn chick results in a decline of serum immunoglobulin and decreased humoral antibody responses. However, chickens treated in this way can still reject tissues from unrelated donors.5, 6 and 7 On the other hand, removal of the thymus from newborn chickens results in defective cell-mediated responses, and in these cases, the manifestations of delayed hypersensitivity are strikingly decreased.5, 6, 7, 8, 9 and 10 In mammals, there also appears to be two functionally distinguishable lymphoid systems; the thymus is evident but the bursa is not a distinct organ, although its functions are thought to be carried out by bursal-type lymphoid cells which are dispersed through the lymphoid system and may be localized in the tonsils and Peyer’s patches. In humans, patients who are athymic display decreased cell-mediated immunity11, 12, 13 and 14 but can produce normal antibody, and there are instances of congenital agammaglobulinemia in which bursal function is lacking.
It is of interest to note that the role of lymphocytes in immunity was unclear until the late 1950s. In the early 60s it became clear that lymphocytes were genuinely immunocompetent. This was based on two principal observations: a) Lymphocytes could transfer cellular and humoral immunity to irradiated rats (which were incapable of giving an immune response themselves); and b) Prolonged drainage of the thoracic duct, which contained large numbers of lymphocytes, impaired the immune responsiveness of the animal.15 In the bird, hemopoietic stem cells from the fetal yolk sac and liver migrate to the embryonic thymus or the bursa, and differentiation to lymphocytes occurs at both of these sites.16 Lymphocytes are seeded from the thymus to peripheral lymphoid tissues and these are referred to as “T” lymphocytes.9,17 Much of this seeding occurs very shortly after birth and so, if the thymus is removed at this time, the animal when mature will have a deficiency of T cells and will exhibit impaired cell-mediated immunity. In the adult, additional stem cells (arising in the bone marrow) migrate to the thymus, and lymphocytes from the thymus continue to be distributed to the peripheral lymphoid tissues. In the bursa, lymphocytes also undergo a “maturation” process and acquire immunoglobulin molecules on their surfaces.18 Seeding of these lymphocytes, which are referred to as “B” cells, to the peripheral lymphoid tissue also occurs. Removal of the bursa results in a deficiency of antibody production.19,20 In mammals, there is no anatomical equivalent of the bursa, but it is thought that the immunologic equivalent of this avian organ lies in the Peyer’s patches and other gut-associated lymphoid tissue. The process of conversion of a hematopoietic stem cell to an immunocompetent cell (either “T” or “B”) occurs in these primary lymphoid areas (thymus or bursal-equivalent tissue) and is independent of antigen-exposure. The conversion of an immunocompetent cell to an effector cell active in cell-mediated immunity or to a cell capable of producing antibody occurs in secondary or peripheral lymphoid tissues and is induced by antigen.
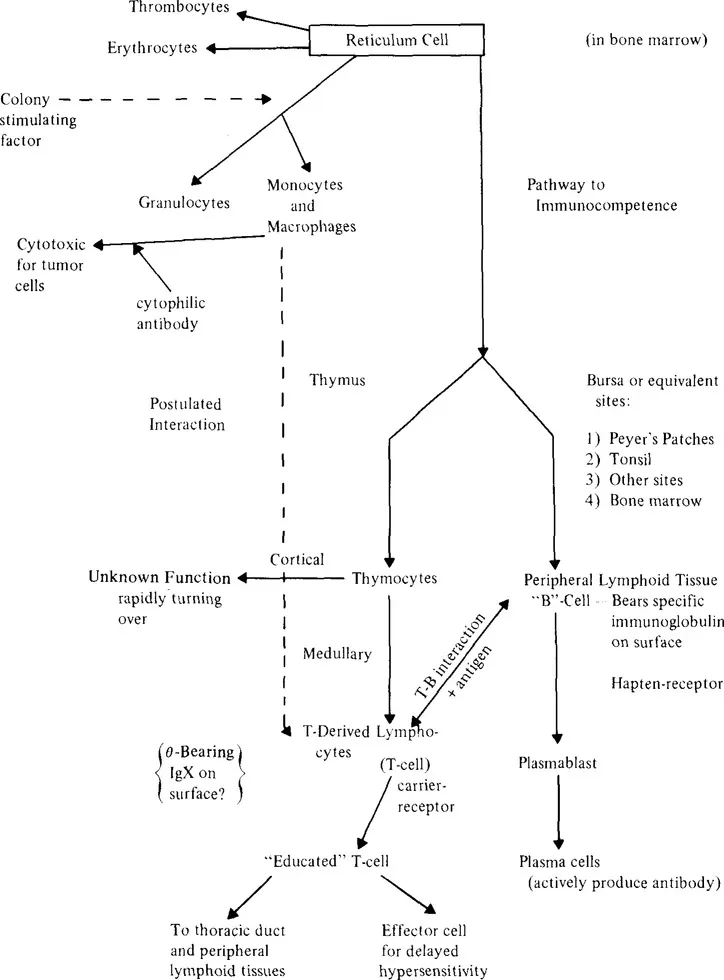
FIGURE 1. The immune system.
In summary, there appear to be two differentiation pathways for cells that react with antigen. A class of lymphocytes migrates from the bone marrow to the thymus where these cells acquire or become capable of expressing an ability to respond to antigen through cell-mediated immunity. These cells are referred to as “T” cells (they should more properly be called thymus-influenced). They mediate the reactions of cell-mediated immunity, as well as graft vs. host (GVH) and homograft reactions. A second type of lymphocyte also arises in the bone marrow and settles in distinct sites in peripheral lymphoid tissue, and in birds, in the Bursa of Fabricus where they give rise to “B” cells.
The ability of a “B” cell which has been exposed to antigen to begin antibody production depends for many, but not all, antigens on concomitant activity of specifically stimulated T cells. These “T” cells may exert an influence on several B cell processes: 1) The switch from IgM to IgG*; 2) Selection of cells producing antibodies of increased affinity; 3) Suppression of “B” cell function under certain circumstances. The “T” cells do not secrete antibody in response to antigen. When a “T” cell is exposed to antigen it can undergo clonal expansion or produce specific products which influence the action of “B” cells, or it can become activated to perform the function(s) of cell-mediated immunity. In the former case, it is common to refer to the “T” cells as being “educated” while in the latter we call the “T” cells “activated.”
Clonal Selection
It is now widely accepted that in the process of differentiation, lymphocytes acquire the ability to react to only one antigen and, in fact, to singular portions of that antigen. This view is formalized in the th...
Table of contents
- Cover
- Half Page
- Title Page
- Copyright Page
- Table of Contents
- Chapter 1 Interactions Between Lymphoid Cells
- Chapter 2 Role of Macrophages in the Immune Response
- Chapter 3 Eosinophils in Humoral and Cell-Mediated Responses
- Chapter 4 The Immunobiology of Tolerance to a Bacterial Antigen: Cellular and Molecular Aspects
- Chapter 5 Molecular Analysis of Lymphocyte Transformation
- Chapter 6 Transplantation: Immunogenetics and Effector Mechanisms
- Chapter 7 The Lung as an Organ of the Lymphoid System
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Developments in Lymphoid Cell Biology by A. Arthur Gottlieb in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over one million books available in our catalogue for you to explore.