Although books exist on the evolution of aging, this is the first book written from the perspective of again as an adaptive program. It offers an insight into the implications of research on aging genetics, The author proposes the Demographic Theory of Senescence, whereby aging has been affirmatively selected because it levels the death rate over time helping stabilize population dynamics and prevent extinctions.

eBook - ePub
Aging is a Group-Selected Adaptation
Theory, Evidence, and Medical Implications
- 243 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
GeriatricsIndex
Biological Sciences1
CHAPTER
Hormesis
Hormesis is where it began for me, though I did not yet know the word. In 1996, I learned for the first time [1] that many animals live longer when they are underfed and their lifespans continue to increase right up to the threshold of starvation.
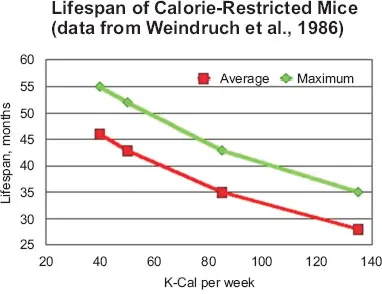
Lifespan plotted against percent CR, based on data from Weindruch and Walford.
This alone is sufficient to imply that aging is an adaptive evolutionary program. Since the Caloric Restriction (CR) phenomenon is reproducible robustly across widely-separated taxa, it must have a general evolutionary basis, a benefit that applies as well to spiders and flies, to dogs and mice, to crustaceans, yeast cells, and worms. But how are they able to extend their lives under the duress of starvation? From what reserve of strength do they pull this ability when starved? And why do they not perform the same trick when they are not starved? All these animals cannot be lengthening their lives when starving unless they hold some capacity in reserve, shortening their lives when they have plenty to eat. Even when they are slightly underfed, they must be holding something back, to be applied only when they are severely underfed. And of course, the feat of maximum life extension is performed under circumstances when the resources available are barely enough for survival.
This behavior contradicts expectations from the “damage” theories that dominate gerontology [2] and the tradeoff theories that dominate evolutionary theory of aging. Most popular and most inconsistent with CR [3] is the Disposable Soma theory [4], based on the idea that the underlying cause of aging is that the body short-changes the calorie budget for repair and maintenance in order to goose the budget for survival and reproduction. If longevity were a function of sufficient caloric energy, then lifespan would not be maximized when consumption is minimized.
When they have enough to eat, animal metabolisms are not trying to live as long as possible. They have evolved to have a shorter lifespan than the maximum of which they are capable. Lifespan is an evolutionary program.
The adaptive benefit of this program is demographic, as articulated by Harrison and Archer [5-7]. The ability to toughen up and to extend lifespan when starved helps the population to survive a famine and delivers the survivors into a world with plentiful resources and thinned competition, in which their progeny may prosper.
When the comparison is made in this way—as extended life during a famine—it appears that there is a benefit for the community and also for the individual. The community resists extinction from famine; the individual delivers its genes into a world where much of the competition has been eliminated, with the opportunity to found a new community. But suppose we frame the same statement conversely: what is the advantage to shortening lifespan when fully fed? This makes it clear that the benefit is only to the community and not to the individual. The individual that dies early just because it has enough to eat does not secure thereby a selective advantage—quite the contrary; dying early carries a cost*. But there is a communal benefit from death on a schedule. If lifespan were indeterminate and open-ended, awaiting a cause of death sufficient to knock out a robust individual in the prime of life, then deaths would be dangerously clumped in time. When conditions were good, almost no one would die; when there was a famine or an epidemic, everyone would die at once. Aging puts death on an individual schedule, so deaths can be spread out through time. Without aging, common sources of death are tightly correlated within a local community, so population swings wildly up and down. Ecosystems are routinely stretched to their demographic extremes and each boom and subsequent population crash poses a risk of extinction.
The evolutionary meaning of fixed individual lifespan is to promote population homeostasis, to make possible stable ecosystems, to avoid population overshoot that risks extinction. In order to have some extra slack to extend lifespan when food is scarce, it is logically necessary to keep lifespan shorter when food is plentiful. You could say that nature invented aging so she would have some room to relent and extend life in times of famine.
This is the thesis on which the remainder of this volume is an elaboration.
A Note on the Use of Teleological Language
“Nature invented aging so she would have some room to relent and extend life in times of famine.”
By convention, biologists avoid this kind of sentence construction. There are no purposes in evolution, just blind mutation and differences between what survives and what does not. To write in terms of purposes encourages a kind of sloppy thinking, with which we may fool ourselves about what may evolve by natural selection and what may not. To be really accurate, what we ought to write instead is:
“Plasticity of mortality under genetic control is an adaptation that aids in the prevention of population overshoot. Populations in which the death rate is left to fluctuate widely with seasonal and stochastic changes in the environment have suffered extinctions, effectively selecting those groups in which there was an adaptive death rate. Plasticity of aging implies higher death rates in the absence of stress, and although this imposes a cost against individual fitness, it delivers a much larger communal benefit through avoidance of extinctions.”
The plausibility of the hypothesis, stated in this way, can only be established by quantitative models, using explicit assumptions about the selective mechanism. This I have done [8-11] and these models will be fully described in Chapter 7.
However, elsewhere in this book, I will honor convention in the breach, freely deploying teleological language and anthropomorphizing nature. I think that the language of purpose helps us to gain intuitive understanding. In practice, every working biologist thinks in terms of purpose and function. The leaf grows toward the light. The sperm swims upstream, seeking the egg. The human metabolism is regulated by a network of chemical signals that makes decisions and transmits message to each cell in the body, to carry them out. Coded in those messages are meanings. It would be absurd to describe biology without this level of understanding.
I was trained in another discipline, physics, in which there is no such proscription. We frequently deploy language such as the following:
“The electron wants to live as near the nucleus as it can get.”
“The photon searches the entire space of pathways from A to B and chooses the one for which the integrated action is a minimum.”
“The crystal seeks its lowest energy state as the temperature is gradually lowered, but may become trapped in an elevated valley, separated from the ground state by a hill.”
Physicists understand that such statements are powerful aids to intuitive understanding, though of course they are no substitute for a quantitative, predictive theory. No one makes the mistake of imagining there is a little man who lives inside the electron and tells it where to go.
In this book, as an experiment, I will defy the convention unapologetically, with a promiscuous use of teleologic language. I will write about evolution with the language of purpose, using metaphors of human intent. I ask the reader to accept this as a shorthand for quantitative, predictive models that will be described in appropriate detail in due course.
The thesis of this book is that:
“Aging and the regulated timing of death evolved for the purpose of stabilizing ecosystems.”
What is Hormesis?
Many homeostatic systems (living and non-living) respond to a disturbance in such a way as to lessen its impact. This is the chemical principle of Le Chatelier [12]. It is a basic property of equilibrium dynamics. When a system is in equilibrium and you push it left, it comes back part-way to the right. It settles in a position further left than originally, but not as far left as you had pushed it. In a homeostatic chemical or physical system that includes a component C, when some of C is removed, the equilibrium shifts in such a way that C is partially restored. The amount of C will still be less than the system had originally, but more than there was after the system was disturbed by removing some C. Conversely, if some C is added, the equilibrium shifts the other way, so that some of the additional C is used up. For example, a buffered solution may have a pH of 7. If you add acid with a pH of 2, it will become only slightly more acidic, perhaps 6.5.
We expect that if there is a shortage of resource X in the environment, an animal or plant, evolved for a plastic adaptive response, adjusts its metabolism so as to mitigate its loss. It adapts by shifting strategies or by substitution, so that it uses less of X. Thus, despite the condition of scarcity, it does almost as well with less X than with sufficient X.
Hormesis is far stranger than this. Animals (and sometimes plants) over-compensate to a stressor, such that they actually do better with stress than without. They live longer when stressed compared to the unstressed lifespan. When X is scare, the animal overcompensates and lives longer than when X was plentiful. When poisoned with toxin Y, the animal overcompensates and actually lives longer than if it were not poisoned.
Hormesis is a property of biological systems, with no analog in physical systems. Compensation is a natural result of homeostasis, but over-compensation is an active process. Hormesis is an evolved system for actively maintaining homeostasis.
Without aging, death would only be from external causes. These tend to be tightly clustered in time. When there is sufficient food in the environment, no one is dying of starvation; in a famine, everyone dies at once. Aging evolved as a way to take control of the death rate from within the genome. We die one by one on an individual schedule to avoid the eventuality that we might all die at once in a famine. When food is scarce, there is plenty of death from starvation and no need for additional deaths from aging. But when food is plentiful, there is no death from starvation and the death toll from aging rises to take up the slack. Hormesis is an adaptation for leveling the death rate in good times and bad.
Implicit in this logic is that the organism is evolved explicitly to shorten lifespan in the absence of stress.
Examples of Hormesis
An Overview of Caloric Restriction
The connection between less food and longer life predates modern scientific study. Hippocrates hints at it [13]. In 15th-century Venice, Luigi Cornaro wrote a volume titled Discorsi della vita sobria (Discourses on the Temperate Life) about his personal experiments with caloric restriction, supplemented by half a litre of wine daily. Cornaro lived to 102. Benjamin Franklin wrote in Poor Richard’s Almanac (1737, while he was still a skinny youth), “To lengthen thy life, lessen thy meals.” (Franklin grew portly and died of pleurisy, an inflammatory disease, at age 84).
In the depression of the 1930s, the issue of widespread malnutrition was discussed in America. How would it affect people’s health and longevity if they did not have enough to eat? Would children’s growth be stunted permanently? Clive McCay was a young researcher at ...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Table of Contents
- Preface
- Acknowledgments
- Foreword by Michael Rosenzweig
- Introduction
- 1. Hormesis
- 2. The Diverse Demography of Aging
- 3. Aging is not Caused by Accumulated Damage
- 4. Three Prevailing Theories and their Failings
- 5. Aging is Very Old
- 6. Evolving Population Control
- 7. How Might Aging Have Evolved as an Adaptation?
- 8. Reforming Evolutionary Theory
- 9. A New Paradigm for Medical Research
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Aging is a Group-Selected Adaptation by Joshua Mitteldorf in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Geriatrics. We have over 1.5 million books available in our catalogue for you to explore.