![]()
1 Tropical Forests
In tropical Australia (Figure 1.1), the rainforest flora contains a unique association of species, descended from ancestors which became established during the Late Cretaceous/Early Tertiary and were later modified by subsequent climatic changes and by the northward drift of the Australian tectonic plate (Morley, 2000). The flora is mostly autochthonous (with a characteristically Australian sclerophyll element) as dictated by the presumed wide separation of the Australian and south-east Asian tectonic plates before the latter part of the Tertiary (Hill, 1994). These factors have resulted in the present-day unique character of Australia’s tropical forest flora with its high concentration of primitive angiosperms (Barlow & Hyland, 1988), and a variety of arborescent and climbing palms, ferns and epiphytes including many endemic species. Many genera are limited to a single representative. They are all the survivors of the extensive drying that occurred during the middle and late Miocene and the more recent Pliocene (Morley, 2000).
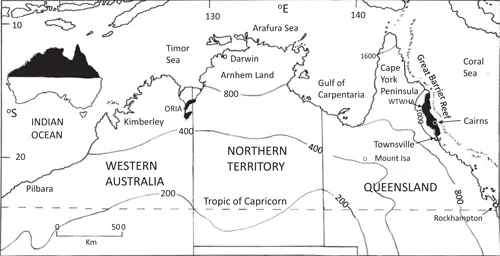
FIGURE 1.1 The tropical north of Australia, showing major locations and bioregions referred to in the text. The annual rainfall isohyets (mm) are estimations based on Australian Bureau of Meteorology data averaged over the period 2000 to 2010. WTWHA, Wet Tropics World Heritage Area; ORIA, Ord River Irrigation Area.
Their species richness, with no clearly dominant species, has been interpreted as reflecting the major crises they have experienced since their first appearance (~75 × 106 years BP) including the meteorite impacts of the Cretaceous, the Eocene cooling, the Late Neogene expansion of the polar ice caps as well as the tectonic movements of the Late Tertiary. They have many features that distinguish them from other tropical forests of the world such as those of the neotropics (South America, the West Indies, Central America south of Mexico), tropical Africa and even the more adjacent Asia-Pacific region.
The so-called wet tropics areas occupy regions experiencing annual rainfalls of between 1,000 and 1,600 mm. The principal plant families represented in today’s tropical rain forests of northern Australia are the Myrtaceae, Lauraceae, Elaeocarpaceae, Rutaceae and Proteaceae. It has been suggested (e.g. Burbidge, 1960) that the present composition of the Australian flora is primarily due to climate selection both within the region and from biotypes available as a result of migration (by communities rather than by chance dispersal of individuals). Most of these tree families are endemic to Australia and only a small number of species extend beyond continental Australia although they may belong to (or have allies in) various regions of south-east Asia (Francis, 1981). It is deduced from geological evidence that rainforest covered much of northern Australia during the early to middle Miocene (23–15 × 106 years BP) but has since that time become restricted to the present-day remnants remaining after the extensive drying of the continent (Long et al., 2002). The fossil record shows little evidence for a post-Miocene influx of Asian plants into Australian rainforests (Ashton, 2003).
These forests have in the past contributed timber products and cabinet woods of great economic value, but a large area, extending as a narrow coastal strip (~100 km wide and 400 km in length) along the eastern side of Cape York is now designated as the Wet Tropics World Heritage Area and protected from further exploitation. Despite its comparatively small size (8,940 km2) relative to other tropical rainforest regions of the world, it is important not only for its conservation value as a remnant of what was a unique and widespread ecological biome but also for its status as a carefully managed and protected area relatively free of the economic and social pressures that threaten rainforests elsewhere (Primack & Corlett, 2005).
Wet Tropics World Heritage Area (WTWHA)
Australia became a signatory of the World Heritage Convention in 1974 and by 2007 there were 17 Australian properties listed. One of them, the Wet Tropics of Queensland World Heritage Area, was listed in 1988. The listing was a recognition of its outstanding natural values including its importance to an understanding of the evolution of the world’s ecosystems as influenced by geological, geomorphic and physiographic features. It was also a response to current reports (e.g. Tracey, 1982) that approximately one-quarter of the total area of rainforest of northern Queensland had been cleared during the 20th century, mainly for intensive production, grazing and agriculture.
The WTWHA (Figure 1.2) is managed by the Wet Tropics Management Authority, answerable to both the Australian Government and the Queensland State Government but working in partnership with other bodies responsible for conservation, research, community engagement and Aboriginal involvement. Day-to-day maintenance of the area is carried out by the appropriate land managers including the State Environmental Protection Agency, Department of Natural Resources and various local government authorities (Australian Wet Tropics World Heritage Authority, 2007–2008).
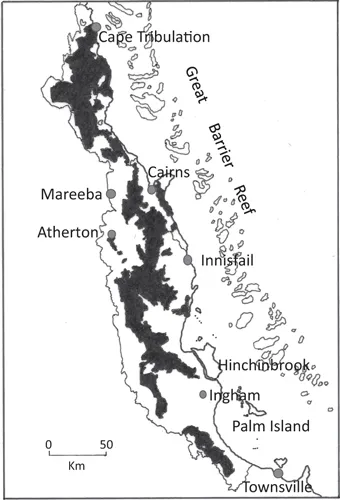
FIGURE 1.2 Wet Tropics World Heritage Area (shaded) extending from 15°26′ S in the north to just north of Townsville (19°16′ S) in the south. (A Directory of Important Wetlands in Australia, 2001, 3rd edition, as published in the Annual Report and State of the Wet Tropics 2010–2011 and reproduced here under the Creative Commons Australia Attribution Licence [CC BY], http://creativecommons.org/licenses/by/au.)
Management of the WTWHA recognises different zones established according to ecosystem integrity, its remoteness from disturbance, the intended physical and social setting, and management purpose. The least pristine zone defined as “compromised forest” is that upon which developed facilities (such as visitor facilities) are allowed. This zone might also include land upon which logging has in the past occurred (before 1988, when commercial logging ceased). Reclassification of such land would depend on the extent of its recovery. The WTWHA has been certified as a tourist destination (Esparon et al., 2013) and is recognised as such by a range of interested parties including tourist operators, environmentalists and volunteer groups.
The combination of high topographic, edaphic and climatic variability within the WTWHA is reflected in a very complex mosaic of different forest types. This has allowed development of models to describe different types of forest growth in terms of the prevailing physiological and physical processes. Two such models, 3-PG (a forest-growth model based on site and climate data) and 3-PGS (driven primarily by satellite data of PAR (photosynthetically active radiation), have been proposed by Nightingale et al. (2008). These models, calibrated and validated against field measurements, contribute to a better understanding of the carbon dynamics of Australia’s tropical rainforest bioregions.
Biodiversity of Tropical Forest Flora
A detailed floristic study of a 25 ha monitoring plot of rainforest situated west of Atherton (17°07′ S, 145°37′ E) in the Wet Tropics bioregion of northern Australia (Sattler & Williams, 1999) was conducted by Bradford et al. (2014). Their initial census of plants having diameters at breast height (dbh) 10 cm recorded 208 species, 128 genera from 53 families with Lauracaea, Rutaceae, Proteaceae and Elaeocarpaceae as dominant. About 83% of the plants recorded were endemic to Australia and 45.2% were endemic to the Wet Tropics bioregion. The data collected provided a baseline description of the rainforest flora against which changes in floristic and stand structure could be measured.
The vegetation of this sample plot was described as complex mesophyll vine forest on granite and meta-sediment alluvium with moderately low soil fertility. The climate of the region is seasonal with 61% of annual rainfall occurring between January and March (mean annual rainfall ~1597 mm). The floristic and structural analysis of the recorded data was described as reflecting the unique nature of Australian wet tropical forests with their high species endemism and affiliations with both Indo-Malayan and Gondwanan flora.
One of the main climatic events affecting the forests of the wet tropics is that due to cyclones, whose frequency and severity are expected to increase as part of anticipated global climatic changes. Studies carried out at the Australian Canopy Crane Research Facility at Cape Tribulation (Figure 1.2) (Stork et al., 2007) have compared the floristic structure of a 0.95 ha lowland tropical forest plot over a 5-year period following the passage of Tropical Cyclone Roma through the area in February 1999 (Laidlow et al., 2007). A survey of the crane plot in 2005 examined 680 stems (≥10 cm dbh) of 82 tree species and showed a post-cyclone increase of 30% in number of stems and a 16% increase in number of species falling into this category. This was interpreted to indicate that the crane site had undergone an active period of post-cyclone recovery suggesting a high degree of temporal stability within the plot despite suffering frequent catastrophic disturbance.
Comparison of the floristic make-up of the crane plot with that of other lowland plots nearby and with mid-elevation plots elsewhere in the north Queensland region showed that the Cape Tribulation site (with Cleistanthus myrianthus, Alstonia scholaris, Myristica insipida, Normanbya normanbyi and Rockinghamia augustifolia as the most abundant species) supports a floristically unique community. There was also considerable variation between rainforest communities across the north Queensland region reflecting differences in local topography, environment and disturbance history.
Today’s wet tropics forest remnants of north-eastern Australia are threatened by a crisis that has the potential to be as destructive as any of those experienced in the past. In addition to the immediate economic consequences of the loss of a range of plant and animal products, the loss of biodiversity, already evident in temperate regions to the south, would impact particularly upon the special role of tropical forests as the primary gene pool from which most temperate plant taxa are thought to have evolved (Bews, 1927). The fossil record shows that the rates and levels of forest destruction now feared, especially if compounded by the effects of climate change, would take many millions of years for full recovery (Morley, 2000).
Forest diversity can be expressed in terms of taxonomic composition, functional and spatial features (including canopy stratification) and the physiological characteristics of the constituent taxa. All of these will influence how the forest responds to climatic or other changes. Fast-growing tree species, for example, may respond more readily to elevated CO2 concentrations than slow-growing species. Equally, species growing under limited nutrient supply may respond differently to CO2 enrichment compared with species under unlimited nutrient supply. For these reasons, investigations of the responses to climate change are best applied under conditions of minimal interference (i.e. growth conditions that are as natural as possible).
The high diversity of rainforest flora, often with the higher taxonomic groups represented by one or a few species, brings particular threats to survival, especially in times of change. Idiospermum australiense, a rainforest canopy tree restricted to a few small populations in north-east Queensland, is the only southern hemisphere representative of the Calycanthaceae. The flowers are protogynous, with some populations being andromonoecious whilst others are hermaphrodite. This species is insect pollinated during its 10–16 day floral life span (Worboys & Jackes, 2005), and although pollen production is large, it is likely that spatial and temporal separation of male and female functions may result in greater vulnerability to climatic change.
Species occupying a specialist niche in the rainforest ecosystem may be particularly vulnerable. Examples are provided by those cases where the life history of the plant and that of an associated animal have become closely linked through a process of co-evolution, as appears to be the case with the rare rainforest tree Ryparosa sp. (Achariaceae) and its associated fruit-eating, ground dwelling avian cassowary (Casuarius casuarius johnsonii). Studies at the rainforest of the Daintree National Park of far north Queensland by Webber and Woodrow (2004) have shown that whilst the young fruit of the Ryparosa have a high concentration of cyanogens, the ripe fruit (consumed by the cassowary) have negligible cyanogen content. The authors further report that passage through the cassowary digestive system improved the germination rate of the Ryparosa seed from 4% to 92% with a much reduced susceptibility of the seed to fruit-fly larvae. Clearly, the intricate balance between the plant and its associated frugivore is crucial to the survival of both partners and the ecosystem.
Observations from other tropical forests of the world (e.g. Laurance et al., 2004) have noted that while tree mortality, recruitment rates and growth have increased over recent decades, there has been a decline in the dominance or density of the slower-growing trees, including species occupying the sub-canopy. Such compositional changes might be expected to impact upon the capacity for carbon storage (see later) and, of course, upon the other forest biota. The extent to which these changes are linked to climate changes is the subject of considerable current speculation.
Any significant loss of biodiversity is known to affect the integrity and functioning of forest ecosystems (Scherer-Lorenzen et al., 2005) confirming observations from early forest research that the yield from mixed forests (on good soils) can, on average, be up to 10–20% (or even up to 50%) higher than that from monoculture in the same area (Kenk,...