![]()
chapter one
Introduction to plant specialized metabolism
Gen-ichiro Arimura and Massimo Maffei
Contents
1.1 Overview
1.2 Molecular evolution
1.3 Human health needs
References
1.1 Overview
Before focusing on the topic of plant specialized metabolites (PSMs, traditionally referred to as plant secondary metabolites), we need to make a general distinction between metabolic pathways essential for growth and development (i.e., primary metabolism) and those not essential for such purposes (specialized metabolism). The availability of carbon, nitrogen, and sulfur resources has a major impact on the production of specific classes of primary metabolites and consequently on the levels and composition of PSMs derived from these primary metabolites (Aharoni and Galili 2011). The focal point in framing the PSMs in a metabolic and functional context is to define their “dispensability” or “indispensability.” The dispensability of these molecules in the face of growth and development has led in the past to consider these molecules as “the ebbs and flows on the metabolism beach” or even waste and/or detoxification products. Today we know that many PSMs are used as chemical signals in the ecosystem and as tools of metabolic defense. PSMs may therefore be regarded as molecules that are “dispensable” for growth and development (with the exception of those metabolic pathways involved in the synthesis of plant hormones), but “indispensable” for survival of the species. Many times the difference between the primary and specialized metabolism is best expressed in functional rather than structural terms, since the same compound can exhibit qualities of both primary and specialized metabolites (Harborne 1993). The most evident qualities of PSMs are definitely their enormous structural diversity, the restriction of their presence to certain families or genera, and their high interspecific and intraspecific variability.
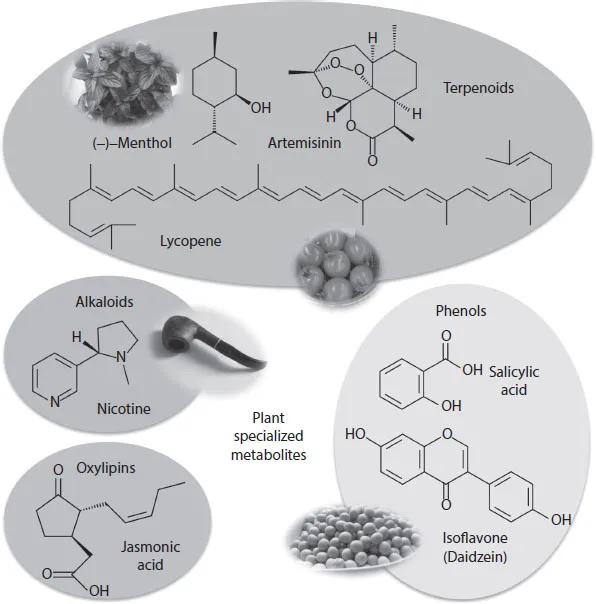
In the genomics age, various genomic tools have transformed the study of PSMs. The field of metabolomics is continually adding novelty and complexity to our information on the chemistry of PSMs, and the availability of whole-genome sequences for an ever-increasing list of plants enables us to examine the genomic basis of PSM production (Zhao et al. 2013). Now just beyond the genomics age, it has become the post-genome era. In recent years, a huge number of genes and gene products involved in PSM metabolism (~200,000 metabolites) have been discovered and characterized. PSMs that are known terpenoids, phenols, alkaloids, fatty acids, and so forth, play several important roles in plant growth or development, environmental adaptation, and trade and defense toward other surrounding organisms (Figure 1.1). Fraenkel (1959) classically stated, “The occurrence of PSMs is sporadic but may be specific for a narrow set of species within a phylogenetic group. It is almost inconceivable that PSMs play a function in the basic metabolism of plants but they significantly function in repelling and attracting insects.” Notably, recent biological and biochemical research involving metabolomic analysis have awakened us to the importance of PSMs, while also disclosing the sophisticated mechanisms underlying plant life cycles and interactions. For instance, plant-derived volatile organic chemicals (e.g., terpenoids and green leaf volatiles) mediate a web of communications between plants and their animal partners, including pollinators, herbivores, and herbivore enemies, therefore contributing to the configuration of biological habitats, that is, ecosystems, and those eventually lead to the diversity and coevolution of organisms. Therefore, the old concept that PSMs have minor functions as “secondary metabolites” is no longer accepted.
Figure 1.1 (See color insert.) Representative plant specialized metabolites.
1.2 Molecular evolution
The great breadth of PSMs is not a coincidence, rather it is the result of a harmonious adjustment of various biogenetic pathways that have been fully integrated into the primary metabolism during molecular evolution. According to several authors of this book, it appears evident as to how plants use the limited numbers of basic metabolic pathways from which diverge an infinite number of variations that lead to hundreds of thousands of molecules. One possible explanation of the origin of PSMs can be sought right in the primary metabolism: some intermediates or side compounds of a primary metabolic pathway are accidently produced due to metabolic impairments caused by environmental pressure. The biosynthesis of such primary metabolites can create additive metabolic flow(s) by a chain of “assembly” that leads to catabolic or anabolic end products. Sometimes such a new metabolic flow triggers a feedback or competitive reaction that acts on the origin of primary metabolism either positively or negatively. However, if excess primary metabolite is available for a parallel metabolic pathway to form a characteristic PSM(s), then the pathway may play an important role in the homeostasis of the primary metabolism. Similarly, several other “parallel” metabolic pathways produce arrays of specialized metabolites that eventually give rise to the hundreds of thousands of PSMs.
One of the characteristics of PSMs is the high “degree of freedom” of compounds, which can vary in amount (concentration) or in quality (chemical structure) irrespective of the development and growth of the organ where they are produced. This is different from the case of primary metabolism, in which all the components required for plant growth and development processes must be sustained at the appropriate cellular levels for the organism’s structural and functional integrity. An important consequence of the high intraspecific variation of PSMs is the capacity to evolve rapidly. It is becoming increasingly clear that trait variance linked to both macro- and microenvironmental variations can also evolve and respond more strongly to selection than mean trait values. Multiple selection forces acting across many spatial and temporal scales probably maintain PSM polymorphisms, but convincing examples that recognize the diversity of plant population structures are limited (Moore et al. 2014).
Undoubtedly, the transition of plants to the land (terrestrialization) was one of the most significant evolutionary events in the history of life on Earth. Plants made their appearance ~400 million years ago, during the Silurian age. At that time, there were already scorpions and centipedes, and scientists have not ruled out the possibility that during the Ordovician (Early Silurian) the first organisms appeared on land, including arthropods feeding on algae and cyanophytes that were predominant even before the Cambrian. Namely, at that time, plant feeders (ancient herbivore arthropods) may have coexisted with plants. During the Permian, flora and fauna changed slowly, and gymnosperms and cycads took the place of Licopodales and tree ferns. At that time, many insects, probably 50% of the species, fed on plants. Butterflies appeared at the beginning of the Cretaceous period and contributed to the process of pollination of flowering plants, which gradually evolved. Herbivorous insects were dominantly present in the Paleozoic Era, as well as during the Mesozoic era. It is likely that the algae Charoficeae gave rise to the terrestrial plants that later evolved into woody plants. In fact, predation of leaves and seeds had not occurred until the Carboniferous, given that the first documentation of feeding dates back to the Mississippian period and predation of seeds in the early Pennsylvanian. Fossil evidence of other feeding methods in the Paleozoic Era, such as the formation of galls and root herbivory, appears even later, from the middle to the end of the Pennsylvanian (Labandeira and Sepkoski 1993). Clearly, PSMs helped ancient plants to withstand attacks from herbivores, as seen in contemporary ecosystems (see Chapters 9 and 13). According to Theis and Lerdau (2003), the evolution of PSM function in such plant defense against herbivores can occur as a result of change at any of several scales of biological organizations. Changes in the distribution of PSMs within plant taxa and in the structures used for their storage can be involved in functional shifts. Moreover, as described in Chapter 9, in some PSMs such as volatile compounds, evolution of function can occur through changes in biological communications using a vapor of PSMs as airborne signals. A better understanding of the evolution of PSMs requires studying the regulation of function across all of these scales.
The broad range of the chemical diversity and biological activities of PSMs raises many questions about their roles in nature and the specific traits leading to their evolution. The answers to these questions would not only be of fundamental interest but may also provide lessons that could help to improve the screening protocols of pharmaceutical companies and strategies for rational PSM engineering (Jenke-Kodama et al. 2008). Since a given structural type of PSM has almost invariably arisen on a number of occasions in different plant taxa, the co-occurrence of a structural class in two taxa is a possible, but not a conclusive indication of a monophyletic relationship of the need for such a PSM. The co-occurrence of a structural class of PSM in two taxa could be due to either convergent evolution of the responsible gene(s) or variable regulation of the genes: in some cases, the genes encoding the enzymes for the production of PSMs with a given structure or structural skeleton evolved at early stages of plant evolution. Even if these genes are not structurally lost during evolution, their expression may cease suddenly because of some accident in, for example, genetic and epigenetic modification. In some cases, however, those genes can be “switched on” again at some later point, leading to a gap in the molecular evolution of PSMs (Wink 2003).
1.3 Human health needs
PSMs are also highly valuable for human life, being used as medicines, flavorings, self-medication drugs such as dietary supplements, and industrial products such as rubbers made from long carbon chains (Figure 1.1). For instance, isoflavones (flavonoid family), produced by most members of the Fabaceae family, are believed to reduce the risk of hormone-dependent cancers, including breast cancer and endometrial cancer, due to their estrogen-like activity (Eden 2012; Varinska et al. 2015). Soy isoflavones are also widely acknowledged to exert various favorable effects against diseases, especially including prevention of cardiovascular disease, osteoporosis, and adverse postmenopausal symptoms, and improvement of physiological conditions, such as maintaining cognitive function (Ko 2014), although soy products have been reported not to relieve menopausal vasomotor symptoms any more effectively than placebo (Eden 2012). Similarly, a wide array of PSMs has been characterized as healthy compounds. On the other hand, nicotine (alkaloid family) is an addictive substance that can cause not only lung cancer, but can also have deleterious effects on pregnancy outcome and the reproductive health of offspring (Wong et al. 2015). Similarly, a broad range of medical, pharmacological, and biochemical studies have clarified various traits of several PSMs (valuable or not and, if valuable, their solo and additive functions), and recent advances in biotechnology enables the production of PSMs in various more efficient, more economical, and more ecologically friendly ways, in addition to the classically used methods for extraction and purification from either cultivated and natural plants, or for production by industrial chemical reactions.
Today, despite the radical changes enabled by chemical synthesis, large numbers and amounts of drugs used for treating various diseases are extracted from plant resources directly or indirectly. Substances that were once extracted from plants have in many cases given way to synthetic molecules, which can be obtained with more effective and definitely cheaper procedures, but this is not true in all cases. Recently, a number of novel molecules discovered in plants have appeared on the market as medicinal products and established a wave of novel pharmacological and clinical studies and research. In particular, there has been significant progress in using these molecules for treating diseases such as cancer or diseases caused by infectious agents such as malaria. Aside from the pharmaceutical industry, which extracts and purifies bioactive substances from plants, there is a popular industry, less standardized and controlled, which uses medicinal plants in the form of herbs, extracts, and powders for the treatment of a large variety of health disorders (Astin et al. 1998).
Notably, the discovery of natural substances is frequently made based on knowledge of customs and traditions, through ethnopharmacologic and ethnopharmacognostic studies, contributing substantially to innovation of medical products by providing novel chemical structures and mechanisms of action.
References
Aharoni, A. and G. Galili. 2011. Metabolic engineering of the plant primary-secondary metabolism interface. Curr. Opin. Biotechnol. 22: 239–244.
Astin, J.A., A. Marie, K.R. Pelletier, E. Hansen, and W.L. Haskell. 1998. A review of the incorporation of complementary and alternative medicine by mainstream physicians. Arch. Intern. Med. 158: 2303–2310.
Eden, J.A. 2012. Phytoestrogens for menopausal symptoms: A review. Maturitas. 72: 157–159.
Fraenkel, G.S. 1959. The raison d’etre of secondary plant substances; these odd chemicals arose as a means of protecting plants from insects and now guide insects to food. Science. 129: 1466–1470.
Harborne, J.B. 1993. Introduction to Ecological Biochemistry, 4th edition. San Diego, CA: Academic Press.
Jenke-Kodama, H., R. Müller, and E. Dittmann. 2008. Evolutionary mechanisms underlying secondary metabolite diversity. Prog. Drug Res. 65: 119, 121–140.
Ko, K.P. 2014. Isoflavones: Chemistry, analysis, functions and effects on health and cancer. Asian Pac. J. Cancer Prev. 15: 7001–7010.
Labandeira, C.C. and J.J. Sepkoski, Jr. 1993. Insect diversity in the fossil record. Science. 261: 310–315.
Moore, B.D., R.L. Andrew, C. Kulheim, and W.J. Foley. 2014. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 201: 733–750.
Theis, N. and M. Lerdau. 2003. The evolution of function in plant secondary metabolites. Int. J. Plant Sci. 164: S93–S102.
Varinska, L., P. Gal, G. Mojzisova, L. Mirossay, and J. Mojzis. 2015. Soy and breast cancer: Focus on angiogenesis. Int. J. Mol. Sci. 16: 11728–11749.
Wink, M. 2003. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry. 64: 3–19.
Wong, M.K., N.G. Barra, N. Alfaidy, D.B. Hardy, and A.C. Holloway. 2015. Adverse effects of perinatal nicotine exposure on reproductive outcomes. Reproduction. 150: R185–R193.
Zhao, N., G.D. Wang, A. Norris, X.L. Chen, and F. Chen. 2013. Studying plant secondary...