This volume provides an introduction to current research on the relation between brain development and the development of cognitive, linguistic, motor, and emotional behavior. At least two audiences will benefit from this book: psychologists interested in brain development, and neuroscientists interested in behavioral development. Although each chapter is content-oriented, the volume as a whole provides a well integrated summary of the latest findings from developmental behavioral neuroscience.

eBook - ePub
Developmental Behavioral Neuroscience
The Minnesota Symposia on Child Psychology, Volume 24
- 248 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Developmental Behavioral Neuroscience
The Minnesota Symposia on Child Psychology, Volume 24
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
1 Electrophysiologic Indices of Human Brain Maturation and Cognitive Development
Herbert G. Vaughan, Jr.
Diane Kurtzberg
Albert Einstein College of Medicine
Human developmental psychobiology seeks to link emerging psychological and behavioral functions with their underlying neural mechanisms. The development of perception, cognition, emotion and motor performance involves the interaction of environmental experience with the maturing brain (see Greenough, chapter 5). However, maturational changes in the neural substrates that underlie these psychological phenomena are as yet not well defined. There is substantially more information on the facts of psychological development than on their biological underpinnings. Although in principle the relations between brain maturation and psychological development can be addressed at many levels in experimental animals, these investigations are limited in the range of cognitive processes that can be examined. Specifically, language and the cognitive operations it supports can only be studied in humans (see Bellugi, chapter 6).
We have incomplete information on cerebral structure during early human development and data on neuronal interconnections and synaptic density are fragmentary (e.g., Purpura, 1975a, 1975b; Huttenlocher, 1984; Marin-Padilla, 1988). We have even less information on the developmental expression of neurotransmitters and modulators that are thought to control the establishment of neural connections and to mediate the cellular modifications associated with learning (see Parnavelas, Papadopoulos, & Cavanagh, 1988; Johnston, 1988 for reviews).
Studies of human developmental psychobiology must rely primarily on the use of non-invasive techniques to obtain information on brain maturation. It is currently possible to obtain detailed information on the gross anatomical status of brain development using computerized axial tomography and, with finer resolution, through magnetic resonance imaging. Metabolic techniques, such as regional blood flow and positron emission tomography, are invasive and therefore not applicable to normal infants and children. Furthermore, despite the functional localization provided by measures of local cerebral metabolism, these methods are inherently incapable of providing real time information on the neural mechanisms that underlie psychological processes.
In principle, the most valuable information on brain function should be provided by the electrical activity generated by neurons within the functioning brain. In humans, this neural activity is manifested in the electroencephalogram (EEG), and in the electrical signals related to sensory, motor and cognitive processes, called “event related potentials” or ERPs (Vaughan, 1969). ERPs provide unique real time indices of specific brain processes that can be monitored throughout the course of development. In the past, the interpretation of developmental EEG and ERP changes has been hampered by the uncertain neural origins of these scalp-recorded signals. During the past two decades, however, both the brain mechanisms (Vaughan & Arezzo, 1988) and the psychological correlates (Picton & Hillyard, 1988) of ERPs have become increasingly better defined. Although much remains to be learned about the neurophysiological basis and psychophysiological relations of the numerous ERP components that have been identified, these real time physiological indices of specific psychological processes can serve as powerful indices of the psychobiology of human development.
In this chapter we show how ERPs, employed as indices of regional brain maturation and of the spatiotemporal patterns of neural processing within the brain, can begin to elucidate some key aspects of developmental psychobiology in infants and children. Before we consider these possibilities, we will outline the conceptual and empirical framework within which this work is placed. We briefly consider the regional anatomical substrate of brain maturation, and relate that to the neuronal network architecture that underlies brain function.
PRINCIPLES OF BRAIN ORGANIZATION
It is a basic premise of human developmental psychobiology that psychological phenomena are manifestations of brain processes, and that psychological changes during development reflect alterations in brain organization and function. Although the neurobiological substrate of psychological processes is obscure and difficult to delineate in detail, we must nevertheless endeavor to learn as much as possible about human brain maturation in order to understand the neural basis of psychological development.
We take as the starting point for this paper, certain principles of brain organization that guide our research. A central feature of human cerebral organization is its parcellation into regions, each having specific features of intrinsic cellular organization and of connectivity with other brain regions (Braak, 1980; Eccles, 1984; Jones, 1984).
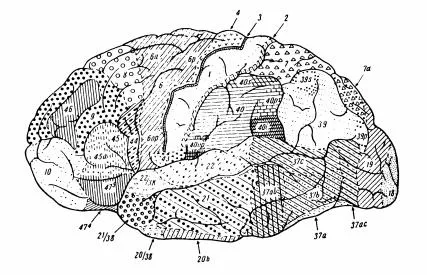
FIG. 1.1 A cytoarchitectonic map of the lateral surface of the human cerebral cortex with areas numbered according to the scheme introduced by Brodman (1909). The cortical areas differ not only in their characteristic laminar patterns of cell body distribution, that are the criterion for the cytoarchitectonic method of parcellation, but also in their patterns of intracortical circuitry and their connectivity with other brain regions. These latter features are the basis for functional differences across cortical regions.
Despite more than a century of controversy, the parcellation of cerebral cortex on the basis of cytoarchitectonics (Fig. 1.1), intrinsic neuronal organization, external connectivity and physiological properties forms the foundation of contemporary cognitive neuroscience (see Peters and Jones, Cerebral Cortex, Vols. 1–7 for detailed reviews of cortical organization). Although there are no isolated “organs of the mind,” there is much to be said for a “modular” approach to understanding the organization of the brain. It is convenient to segregate cortical regions as subcomponents of larger brain systems. Anatomists and physiologists typically select one brain region, and possibly some of the structures with which it connects, for intensive analysis. Nevertheless, even the simplest psychological processes are not limited to a particular patch of brain, as was emphasized by Luria (1973). We accept the idea that psychological processes represent the coordinated interaction of a number of brain regions, each of which has its specialized intrinsic organization and modes of operation (see e.g., Pandya & Yeterian, 1985; Goldman-Rakic, 1988 for further discussion).
A complementary way of looking at brain organization focuses on its network properties. Inasmuch as the brain comprises neurons and their connections, and operates through the activation and modification of these connections, the brain is in fact a neural network. The anatomical organization of these neural networks and the principles that govern modifications in the functional effectiveness of their synaptic connections are key issues to be addressed in order to understand the neurobiological basis of psychological development.
The formulation of “neural network” models has become a prominent area in cognitive neuroscience in recent years, following a long period of development by a small group of pioneering theorists. The basic ideas that underlie contemporary neural network theory were set forth by Donald Hebb in his seminal work The Organization of Behavior (Hebb, 1949). Hebb postulated that brain circuitry comprises two kinds of neuronal networks, which he called “cell assemblies” and “phase sequences.” Cell assemblies are groups of neurons, which through a combination of genetic determinants and repeated exposure to specific external inputs, come to respond to particular stimuli with characteristic, relatively stereotyped patterns of activation. Phase sequences, by contrast, represent more flexible dynamic network activation patterns, comprising linkages of cell assemblies directed by the particular processing demands of the perceptual-cognitive task at hand. Hebb’s theory is a developmental one, in which the establishment of cell assemblies and phase sequences is conditioned by changes in synaptic efficiency, due to repeated associations of a presynaptic input pattern with concurrent activation of the postsynaptic neuron. This associative mechanism for the modification of synaptic efficiency has come to be known as the “Hebbian synapse” and represents a major, although probably not the sole determinant of synaptic changes that occur during learning.
Hebb’s ideas were revolutionary and did not immediately lead to efforts to develop them into formal theory. The work of McCulloch and Pitts (1943) was probably more influential among early mathematical network modelers, including Rosenblatt (1958), whose “perceptron” represented the first attempt to model a specific sensory system, and other workers who incorporated learning into network models. (See Anderson and Rosenfeld, 1989 for a compendium of seminal papers on neural network modeling.)
A competing, and for a time dominant, model of brain mechanisms was proposed by John von Neumann (1958). This model was the serial processing digital computer. At the time, digital computers seemed to provide the nearest analogy to the powerful computational abilities of the human brain. Models of perceptual and cognitive operations that employed serial models similar to the logical structure of digital computers have been especially popular in cognitive psychology, in which the concepts of “processing stages” and “serial search of memory” has been the topic of much experimental work and theory (e.g., Sternberg, 1969). In serial models, memory storage is generally viewed as distinct from the brain structures responsible for sensory, perceptual and cognitive operations. These operations would access memory, much as discrete memory locations are addressed in a computer.
The development of parallel distributed processing (PDP) models, which are essentially Hebbian in their main features, has radically altered our conceptualization of the brain operations that underlie perceptual and cognitive processes (Rumelhart, McClelland et al., 1986; McClelland & Rumelhart et al., 1986). The most important differences include, first, an emphasis on the massive parallelism of brain operations, which is hardly surprising to anyone who knows anything about neuroanatomy; and second, the localization of mnemonic mechanisms within the processing networks themselves, so that memory is an intrinsic property of the network, residing in the efficiency of a synaptic transmission among the neural elements that comprise it. A third key feature of network models is the presence of feedback from later levels of the network to earlier ones. This feature is anatomically embodied in the recurrent projections that are ubiquitous within the brain, and which may form the basis for so-called “top down” processing in cognitive psychology, and “back propagation” in PDP network models. These recurrent connections can modulate the responses of brain regions that process sensory input at earlier stages according to the presumed significance of the input, either filtering out unimportant inputs or selecting information deemed to be of significance for further processing. Recurrent projections can also participate in the operation of “comparator” mechanisms that serve to detect novel or deviant stimulus patterns.
We emphasize that characterization of the brain as a neural network does not imply either that the brain is diffusely organized, with every neuron connected to every other, or that the organization of the networks is similar in all brain regions. Neither of these circumstances is likely to be the case. For example, the manner in which visual and auditory information is processed by the brain differs substantially, as reflected in major differences in the anatomy and physiology of these sensory systems. It is necessary to consider these specialized features of regional brain organization as well as interactions among brain regions when we examine the changes in brain organization that underlie psychological development.
Having briefly mentioned some aspects of contemporary models of brain processes, we now consider how the electrophysiologic study of human brain dynamics may shed light on certain aspects of the development of brain organization.
EVENT RELATED BRAIN POTENTIALS
Event related potentials (ERPs) are manifestations of the activity of large numbers of neurons that are closely synchronized with some externally observable event, such as a stimulus or motor response. ERPs can be extracted from the ongoing spontaneous neuroelectric potentials by averaging a number of EEG samples time-locked to the synchronizing event. The averaged ERPs are enhanced with respect to the amplitude of the random background EEG in proportion to the square root of the number of averaged samples. Thus, even very tiny synchronous brain signals can be resolved, given a sufficiently large number of samples. For example, the extremely small electrical signals generated within the subcortical auditory pathways, which are in the order of tenths of a microvolt at the surface of the scalp, can be recorded by averaging the responses to 1000 or more stimuli. ERPs generated within cerebral cortex, which are usually several microvolts in amplitude at the scalp, can often be satisfactorily recorded with averages of less than 100 trials.
Over the past 2 decades ERPs have been described that represent the cortical and subcortical responses to sensory stimuli, brain activity associated with voluntary movements, and with various cognitive processes (see Picton, 1988 for reviews). The sensory evoked potentials (EPs) represent more or less obligatory responses to the physical properties of the sensory stimuli. Movement related potentials (MRPs) are observed in recordings time-locked to quick extremity or eye movements. MRPs include not only the endogenous brain activity that precedes the movements, but also reafferent activity associated with the movements and changes in sensory input that are a consequence of the movements. A third class of ERPs are those associated with active processing of sensory input and with various cognitive operations. These ERP components are often called “endogenous” potentials to contrast them with the obligatory, “exogenous” EPs. We use the term “processing contingent components” (PCCs), to emphasize that these components represent the processing of sensory input beyond the obligatory responses elicited by the repetitive presentation of a nonsignificant stimulus, and also to distinguish them from MRPs which are also “endogenous.” PCCs are elicited in a variety of tasks requiring active sensory or other cognitive processes, including linguistic processing. PCCs, however, can also be elicited autom...
Table of contents
- Cover
- Half Title
- Full Title
- Copyright
- Contents
- Preface
- 1. Electrophysiologic Indices of Human Brain Maturation and Cognitive Development
- 2. Basic Mechanisms of Human Locomotor Development
- 3. Developing Emotions and Cortical Networks
- 4. Cortical Versus Limbic Immaturity: Relationship to Infantile Amnesia
- 5. Induction of Brain Structure by Experience: Substrates for Cognitive Development
- 6. Language, Cognition, and Brain Organization in a Neurodevelopmental Disorder
- Author Index
- Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Developmental Behavioral Neuroscience by Megan R. Gunnar,Charles A. Nelson in PDF and/or ePUB format, as well as other popular books in Psychology & Cognitive Neuroscience & Neuropsychology. We have over 1.5 million books available in our catalogue for you to explore.