Aggression usually involves a sequence of behaviors, reflecting escalations and de-escalations in the form or intensity of the actions taken, which play out over time. This book provides a context in which social and biological research on the aggressive behaviors of human and non-human subjects, interacting in dyads or groups, can be compared and integrated. Implicit in this juxtaposition is the major question of whether general principles governing the dynamics of aggression within and between episodes may be discerned. Aggressive behavior is described at different levels of analysis in humans and a number of other animal species. Three basic views of aggression dynamics become apparent:
* The economic interpretation: Aggression will be escalated when it pays one of the combatants to do so or, more generally, when the potential benefits outweigh the risks. Decisions to escalate or de-escalate are part of a calculated "strategy", in one or another sense. This interpretation is formalized within game theoretic models as applied to animal conflicts and to international conflicts, within the chapters of this text.
* The psychological process interpretation: Emphasis is placed on psychological/physiological processes within the individual. The chapters stress the importance of acute emotional states of anger and aggressive arousal and argue the role of peripheral sympathetic activation, while proposing a central neural mechanism. Children escalating their tantrums, adult humans and animals of other species intensifying their interpersonal conflicts, national leaders going to a war footing all appear to suffer a narrowing of attention and progressive failure of cognitive function under the intensifying stress of conflict. Perhaps these changes in attention, sensory and cognitive functions, and risk taking reflect a "commitment to aggression" which is necessary for organisms to engage in potentially dangerous and painful encounters.
* The emergent process interpretation: Escalation emerges in a spontaneous and dynamic way as the actions of one participant elicit reactions from the other(s).

eBook - ePub
The Dynamics of Aggression
Biological and Social Processes in Dyads and Groups
- 368 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Dynamics of Aggression
Biological and Social Processes in Dyads and Groups
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
Experimental PsychologyIndex
PsychologyPART I
Differing Perspectives on Aggression Dynamics: Introduction to Game Theoretical Analyses and Arousal Hypotheses
CHAPTER 1
Game Theory Models and Escalation of Animal Fights
University of Central Lanchashire, United Kingdom
University of Glasgow, United Kingdom
ESCALATION IN ANIMAL FIGHTS
Fighting is widespread in the animal kingdom (Archer, 1988; Huntingford & Turner, 1987), occurring most commonly over access to valuable but limited resources, such as food or mates. The exact form of the behavior is very variable; anemones club each other with specialized tentacles, crickets kick each other and lash out with their antennae, frogs wrestle, gulls peck, and gorillas beat their chests. There are, however, some features that are common to the great majority of animal fights, one of these being the occurrence of escalation.
Most species have a number of different aggressive actions in their repertoire, and these are deployed in a characteristic sequence. Fights often begin when the opponents are at a distance from one another, exchanging movements or displays that are of relatively low intensity and therefore low cost both in terms of energy consumption and the likelihood of damage. If these low-intensity actions do not cause one or the other contestant to withdraw, then patterns of behavior that are progressively more energetic and more risky come into play, culminating in the most intense and damaging acts in the animals’ repertoire. These are the most effective in inducing withdrawal or submission in the opponent. This progressive increase in the intensity and cost of behavior as a fight proceeds is referred to as escalation.
Escalation has been described in detail in many ethological studies of animal fights. One example is a laboratory study of the cichlid fish (Nannacara anomala) by Jakobsson, Radesater, and Jarvi (1979), which fight over status within a social group. In the initial phases of the fight, the fish commonly engage in actions such as lateral display and tail beats that are performed at a distance and involve a low risk of injury. As the fight progresses, these actions drop out in favor of mouth wrestling and biting, which involve direct physical contact and are more intense; in this species escalated fighting carries a greater risk of injury, in the form of lost scales and damaged fins.
A second example comes from a field study of iguanas carried out on an islet near Panama by Rand and Rand (1976). Iguanas (Iguana iguana) come here to breed, and the females dig burrows in which they lay their eggs; digging burrows is hard work and occurs over several days, interspersed with resting periods away from the burrow. Disputes arise when a female returning from resting tries to take over a burrow that currently has a resident. Here again, fights involve an escalating series of displays that can be characterized in terms of increasing cost, cost for these ectothermic animals being mainly in terms of energy expenditure. Thus, fights progress from open mouth display to head swing, to huff, lunge, and bite, each one bringing the opponents nearer together, as well as being more energetic and more effective in intimidating a rival.
As a last example, red deer stags (Cervus elephas) fight during the rut over access to females (Clutton-Brock, Guinness, & Albon, 1982). Initially, they simply roar at each other from a distance (Fig 1.1a), and the male with the slowest roar often retires at this point. However, some fights progress to the more intense stage of parallel walking (Fig. 1.1b), and of these a proportion proceed to antler wrestling (Fig. 1.1c), which is energetically costly and may result in injury.
In all three species, fights stop when one animal, the loser, gives up, thus leaving its opponent victorious. The behavior shown by the winner and loser are similar and indicative of aggressive motivation until near the end of the fight; the eventual loser then reverts rapidly to low-cost actions (de-escalation) and withdraws from the encounter. For example, early on in fights between cichlid fish (Nannacara anomala), both the eventual winner and the eventual loser tend to respond to a bite (a high-cost, escalated action) by retaliating in kind. However, just before the end one animal starts responding to a bite with tail beating and frontal displays, and this is the eventual loser (Jakobsson et al., 1979).

FIG. 1.1. Fight between red deer stags during the rut over access to females: (a) roaring from a distance, (b) parallel walking, (c) antler wrestling (after Clutton-Brock et al., 1982).
GAME THEORY AND ANIMAL FIGHTS
Such ethological studies provide detailed descriptions of the process of escalation but do not explain why escalation occurs, either in terms of its evolution or the behavioral mechanisms responsible for it. Recent attempts to explain the evolution of animal fighting make extensive use of a branch of applied mathematics called game theory. These analyses investigate the evolutionary consequences of using different patterns of fighting by depicting animals fighting in these different ways as players in a game. The earliest game theory models of animal fights were used to explain why, given the great rewards for winning, animals do not always fight as fiercely as possible; in other words, they set out to explain the existence of both low-intensity and high-intensity forms of fighting. Later models dealt with, among other things, the question of why animal fights so often take the form of a sucession of increasingly intense and costly actions; in other words, these models explain the occurrence of escalation.
EXPLAINING THE EXISTENCE OF HIGH-AND LOW-INTENSITY FIGHTING
The simplest game theory model of animal fighting is the hawk-dove model (Maynard Smith, 1976, 1982). This model considers two individuals competing for some resource to which one gains exclusive access if its opponent retreats or is injured. The players are identical except that doves use only low-cost displays and continue until one withdraws, whereas hawks always launch straight into fierce, high-cost fighting, stopping only when one animal is injured. In the game theory literature, hawks are sometimes described as showing escalated (as opposed to conventional) fighting, which is different from the ethological use of the term (see Caryl, 1981).
The costs and benefits gained by hawks and doves in fights against opponents using these same two strategies are calculated (based on preset rules that determine the outcome of each kind of fight) to give the payoff matrix shown in Table 1.1. The analysis works out the overall fitness of individuals pursuing each strategy and from this the relative frequency of the strategies in successive generations. In particular, it works out if either strategy is evolutionarily stable, meaning that when it is common it cannot be bettered by any other strategy in the game under the cost-benefit conditions defined by the game’s rules. Evolutionarily stable strategies (ESSs) are how we would expect real animals to behave if the game has defined correctly the costs and benefits of high-risk and low-risk fighting. In this game, hawk is an ESS when the benefits of victory outweigh the costs of injury; when the converse is true, even though hawks always defeat doves, because costs are high they lose out, so that neither is an ESS; there is, however, a certain (evolutionarily stable) proportion of doves and hawks at which the two strategies gain equal payoffs.
TABLE 1.1
Payoff Matrix for the Hawk-Dove Game
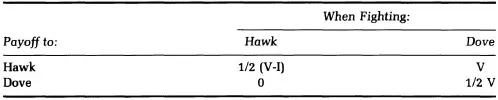
Note. V = Benefit accruing to victor. I = Cost of injury.
PREDICTING THE OCCURRENCE OF ESCALATION
By showing that, because of the costs of injury, hawk is not always an ESS, the hawk-dove model explains the co-existence of high- and low-intensity forms of fighting. However, the simple hawk-dove dichotomy was set up to investigate the fitness-related consequences of using displays rather than overtly aggressive actions and so does not depict the progressive increase in intensity described in the ethological studies of animal fights (Caryl, 1981). Later we show other game theory models that can help us to understand certain features of the process of escalation.
The Importance of Risk of Injury
So far, we have described the process of escalation as if all fights in a particular species proceed in an identical way. There is, however, great variability in whether and to what extent animal conflicts become escalated. In some cases, one animal may flee relatively quickly without engaging in intense, costly fighting; in other encounters involving the same species, and perhaps even the same individuals on a different occasion, fights may be long and fierce. Game theory models can help us to predict the circumstances in which we would expect fights to escalate to high-cost actions by specifying in functional terms when it will no longer be advantageous for an animal to continue fighting. To anticipate and oversimplify, escalated fights are to be expected when the animals concerned are equally matched in fighting ability and when both place a high value on the disputed resource.
The game theory model most relevant to predicting the occurrence of escalated fights is Parker’s (1974) assessment strategy model. Building on Maynard Smith’s earliest analyses (Maynard Smith, 1972; Maynard Smith & Price, 1973), Parker set out the following simple but compelling argument: Any individual that is able to assess how its own fighting ability (designated as resource holding power or RHP) compares with that of its opponent will be at a selective advantage, because it could withdraw without damage when its own RHP was lower but attack (with a good chance of winning) when its own RHP was higher.
The simple concept of RHP enables us broadly to predict the circumstances when escalated fights will occur. These exist when the opponents’ RHPs are reasonably well matched; when they are not, the one with the lower RHP will withdraw after an initial assessment or “sizing up” phase (Parker, 1974). These principles have been incorporated into formal game theory models (Maynard Smith, 1982, Maynard Smith & Parker, 1976), which extend the original hawk-dove game. The additional feature is a strategy called assessor that attacks (plays hawk strategy) if its RHP is higher and displays (plays dove strategy) if it is lower. Where escalation proves to be costly, as in the case of animals with dangerous weapons, the assessor strategy is shown to be an ESS (Table 1.2). More precisely, assessor will be an ESS when the cost of losing an escalated fight is greater than the cost of the assessment phase. Under the opposite conditions, where assessment is costly yet escalation is less dangerous, the hawk strategy of unconditional attack is the ESS.
Assessment may be more costly than escalated fighting in ectothermic animals (i.e., where energy resources can be problematic) without dangerous weapons, for example, in toads and newts (Davies & Halliday, 1978; Smith & Ivins, 1986). However, many kinds of animal have weapons (teeth, claws, hoofs, or horns) that are capable of wounding a rival, and in these cases losing an escalated fight is likely to involve a relatively high cost. Here, therefore, the hawk-dove-assessor model predicts that the assessor strategy is an ESS. In other words, we should expect that such animals will adapt the intensity of their aggressive responses to the relative RHP of a potential opponent. Many studies of animal fights have shown that this is, indeed, the case (Archer, 1988, Table 9.1).
TABLE 1.2
Payoff Matrix for the Hawk-Dove-Assessor Game

Notes. V = Benefit accruing to victor. I = Cost of injury. A = Cost of assessment.
This raises the question of how an animal is able to assess its opponent’s resource holding power–or fighting ability–relative to its own. In practice, a number of cues may be used, but because bigger animals are usually stronger perhaps the most widespread one is the opponent’s size. The sensory channels through which size is assessed must be variable, because the larger of two opponents is more likely to initiate a fight in animals as varied as the sea anemone (Brace & Pavey, 1978), a Gastropod mollusc (Zack, 1975), hermit crabs (Mitchell, 1976), field crickets (Dixon & Cade, 1986), and a wide variety of vertebrates (see Archer, 1988, Table 9.1). Tail beating in male cichlid fish (Nannacara anomala, see earlier) may perhaps create pressure waves whose size depends on the size of the performer; certainly, in red deer only strong males can roar fast (see before), and stags can use this to identify large differences in RHP at an early stage in an encounter.
Knowledge gained about fighting ability from previous encounters will also provide a powerful cue for animals that live in social groups and are able to recognize one another individually (Van Rhijn &Vodegel, 1980). Such learning will form the basis of dominance hierarchies. It has often been noted that recognition of the superior fighting ability of a more dominant animal by a subordinate is crucial for maintaining dominance relationships (Gartlan, 1968; Rowell, 1966; Tinbergen, 1953). In primate societies, where individuals may gang together to defeat a high-ranking opponent, assessment of the probability of winning an encounter will depend not just on the individual RHP and past experience of the main contestants but also on the presence and RHP of potential allies (Datta, 1983).
The Importance of Resource Value
There are, therefore, a wide range of cues an animal might use to assess its opponent’s fighting ability. The empirical evidence generally supports the prediction that animals assess relative RHP and that longer, fiercer fights (escalated fights in game theory terminology) occur between opponents whose fighting abilities are similar. Nevertheless, as Parker (1974) recognized, a model based only on relative RHP is a naive one. Even though the concept of RHP has proved one of the most important insights to come from game theory analyses of animal fights, it is not the whole story. Several other factors must be considered. All game theory models are based on calculations of the payoffs to the participants, given that certain costs and benefits are involved. RHP involves one very important aspect of the costs of fighting, but we have not yet considered the benefits to be obtained from winning a fight.
Most cases of animal fights to which game theory analyses have been applied are competitive in the sense that the protagonists are competing for a resource such as food, a territory, a nest site, or a potential mate (Archer, 1988). When considering the benefits of winning a fight, complications arise if the resource is not of equal value to the two individuals; for example, the same piece of food may be of more value to a starving animal than to a better fed one and, to give a more complicated example, a male that has invested a certain amount of time and energy in fertilizing a female’s eggs has more to gain by winning a fight over access to the female than does a challenger. Theory predicts that animals will be more likely to escalate to high-cost acti...
Table of contents
- Cover Page
- Half Title page
- Title Page
- Copyright Page
- Dedication
- Contents
- Preface
- Part I Differing Perspectives on Aggression Dynamics Introduction to Game Theoretical Analyses and Arousal Hypotheses
- Part II Aggressive State and Trait Behavioral and Physiological Processes Within Individuals
- Part III Aggression Dynamics in Development Interactions Within and Outside the Family
- Part IV Aggression Dynamics in Larger Social and Political Contexts Game Theory Revisited
- Author Index
- Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Dynamics of Aggression by Michael Potegal,John F. Knutson in PDF and/or ePUB format, as well as other popular books in Psychology & Experimental Psychology. We have over 1.5 million books available in our catalogue for you to explore.