1
Cultural Phylogenies and Explanation:
Why Historical Methods Matter
Carl P. Lipo, Michael J. O’Brien, Mark Collard and Stephen J. Shennan
Imagine finding a collection of things and having to figure out how each one got to be the way it is. In addition to learning about, say, the chemical and physical characteristics of the objects, resolving this issue would lead you to ask about their history. Are they related to one another? If so, are they equally related, or are some items related more closely and some more distantly? These are not easy questions to answer, but they are ones with which natural and social scientists wrestle on a regular basis. They also are central to the chapters in this book.
Interest in genealogical, or “phylogenetic,” relationships has a long tradition in the natural sciences. Although efforts at explaining the natural world in phylogenetic terms can be traced to at least 350 B.C. and Aristotle’s Historia Animalium, most of the major steps in developing a means of describing biological organisms in a way that reflects their affinities have occurred in the last 300 years. One of these was the publication in 1735 of Carolus Linnaeus’s Systema Naturae. Linnaeus popularized what has become one of the core ideas of biological phylogenetics, namely that species can be grouped into a hierarchy of progressively more inclusive taxa. Further progress in building a robust set of methods for delineating phylogenetic relationships among organisms came in 1859 with the publication of Charles Darwin’s On the Origin of Species. Although it was not immediately recognized, Darwin’s theory directed biologists interested in relatedness to limit their descriptions to one form of similarity—the kind that results from heredity. Traits that two or more taxa share because they inherited them from a common ancestor are termed “homologies.”
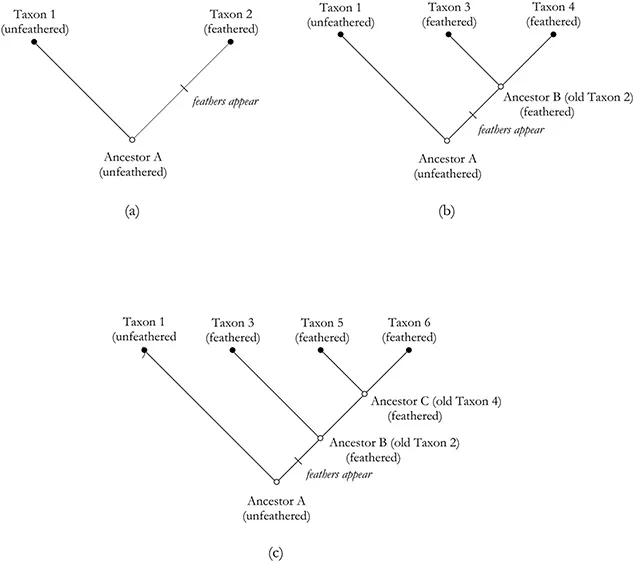
A robust numerical method that made use of this portion of Darwin’s theory emerged in the twentieth century at the hands of Walter Zimmerman (1931) and especially Willi Hennig (1950, 1966). The key component of this method, termed “phylogenetic systematics,” or more commonly “cladistics,” is its focus on a subset of homologous traits, namely those that are considered to be “derived” rather than “ancestral.” Derived traits are character states exhibited by a set of sister taxa and their immediate ancestor but no other taxon (figure 1.1). The final products of cladistic analyses are treelike structures called “phylogenetic trees,” or “cladograms,”1 that depict relationships among taxa.
Figure 1.1
Phylogenetic Trees Showing the Evolution of Six Taxa (after O’Brien et al. 2001)
In (a) feathers appear during the evolution of Taxon 2 out of its ancestral taxon. The state “feathered” is termed an “apomorphy.” In (b) Taxon 2 has produced two taxa, 3 and 4, both of which contain feathered specimens. The appearance of feathers in those sister taxa and in their common ancestor (B) makes it a “shared derived” character state, technically termed a “synapomorphy.” In (c) one of the taxa that appeared in the previous generation (Taxon 4) gives rise to two new taxa, 5 and 6, both of which contain feathered specimens. If we focus attention only on these two new taxa, “feathered” is now an “ancestral” character state, technically termed a “plesiomorphy” (shared ancestral character states are termed “symplesiomorphies”). Note that it is shared by more taxa than just sister taxa 5 and 6 and their immediate common ancestor. But if we include Taxon 3 in our focus, having feathers is a synapomorphy because, following the definition, it occurs only in sister taxa and in their immediate common ancestor.
In Figure 1.1(a) feathers appear during the evolution of Taxon 2 out of its ancestral taxon. The state “feathered” is termed an “apomorphy.” In (b) Taxon 2 has produced two taxa, 3 and 4, both of which contain feathered specimens. The appearance of feathers in those sister taxa and in their common ancestor (B) makes it a “shared derived” character state, technically termed a “synapomorphy.” In (c) one of the taxa that appeared in the previous generation (Taxon 4) gives rise to two new taxa, 5 and 6, both of which contain feathered specimens. If we focus attention only on these two new taxa, “feathered” is now an “ancestral” character state, technically termed a “plesiomorphy” (shared ancestral character states are termed “symplesiomorphies”). Note that it is shared by more taxa than just sister taxa 5 and 6 and their immediate common ancestor. But if we include Taxon 3 in our focus, having feathers is a synapomorphy because, following the definition, it occurs only in sister taxa and in their immediate common ancestor.
The principles that drive phylogenetic methods are not restricted to the study of biological entities. Indeed, phylogenetic methods are simply algorithms for building phylogenies once descriptions of taxa are made. The important point is that phylogenetic methods can be used to relate any set of features that change in nonrandom fashion over time, regardless of the mechanism or process.
In recent years, a growing number of social scientists have begun to use phylogenetic methods, especially cladistics, to address questions of cultural evolution. The datasets used in these studies come from a wide range of locations, including the Pacific (e.g., Gray and Jordan 2000; Hurles et al. 2003; Jordan 1999; Kirch and Green 1992, 2001), Africa (e.g., Foley 1987; Holden 2002; Holden and Mace 1997, 1999; Mace and Pagel 1994), Europe (e.g., Collard and Shennan, 2000; Gray and Atkinson 2003; Renfrew and Boyle 2000), Asia (e.g., Tehrani and Collard 2002), and North America (e.g., Lipo 2001; Lipo et al. 1997; O’Brien and Lyman 2003a; O’Brien et al. 2001, 2002).
The subject matter is similarly diverse. The growing literature on the use of phylogenetic methods in studies of material culture includes applications to stone tools (Foley 1987; Foley and Lahr 1997, 2003; O’Brien and Lyman 2003a, 2003b; O’Brien et al. 2001, 2002; Robson Brown 1995, 1996), baskets (e.g., Jordan and Shennan 2003), pottery (e.g., Collard and Shennan 2000), carpets (e.g., Tehrani and Collard 2002), written texts (e.g., Spencer et al. 2004) and even entire industries (e.g., Anderson 2003). The use of phyloge-netic methods is also seeing a growing usage in sociocultural anthropology (e.g., Borgerhoff Mulder 2001; Borgerhoff Mulder et al. 2001; Holden and Mace 1997, 1999; Jones 2003; Mace and Pagel 1994; Sellen and Mace 1997), in historical linguistics (e.g., Gray and Atkinson 2003; Gray and Jordan 2000; Holden 2002; Jordan 1999; Platnick and Cameron 1977; Rexová et al. 2003), and in multi-disciplinary studies (e.g., Hurles et al. 2003).
Adopting phylogenetic methods that have been developed primarily in biology and paleontology creates a set of theoretical, methodological, and empirical challenges as we attempt to apply the methods anthropologically. From a practical standpoint, do the methods accomplish what we want them to? From an epistemologica! standpoint, are they appropriate methods to use? And at the most fundamental level, are the products something that we as anthropologists need? Put simply, should we even care about phylogeny?
Phylogeny and Cultural Evolutionary Research
Let us return to the example we used in the opening paragraph: we have a collection of things and are interested in figuring out how each got to be the way it is. This is a phylogenetic problem. For the sake of argument, let us say that the things in the collection are artifacts recovered from an archaeological site. One means of creating a sequence is to examine artifact form. Analysis of form has long been used as a means of studying cultural continuity, the assumption being that artifact similarity often is a function of common ancestry (O’Brien and Lyman 1999). As Albert Spaulding (1955: 14) argued, variation in material culture “can be related to the proposition that cultural change is systematic rather than capricious and to the auxiliary proposition that an important basis for the systematic behavior of culture is its continuous transmission through the agency of person to person contact.” Artifact variation, when described appropriately, can be explained as a function of descent with modification. Archaeologists such as A.V. Kidder (1932) and James Ford (1936) knew this, and it formed the basis of the approach that came to be known as culture history (Lyman et al. 1997).
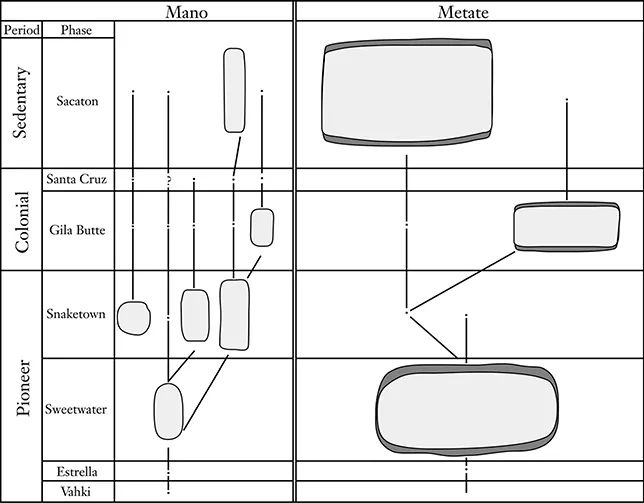
The culture history approach to anthropology is broadly compatible with the biological model that views heritable change as descent with modification. Archaeologists have been modeling change in artifact form in this way since the birth of the discipline (e.g., Evans 1850; Petrie 1899). One example is E. B. Sayles’s (1937) diagram (figure 1.2) showing the evolution of manos and metates from Snaketown, Arizona. In this figure, Sayles makes the claim that mano and metate forms have a single ancestor and diversify over time. This is descent with modification.
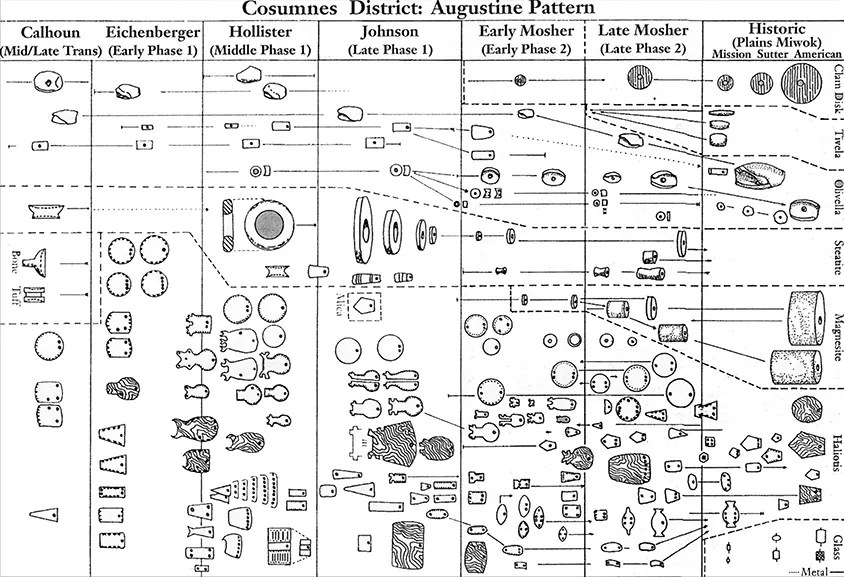
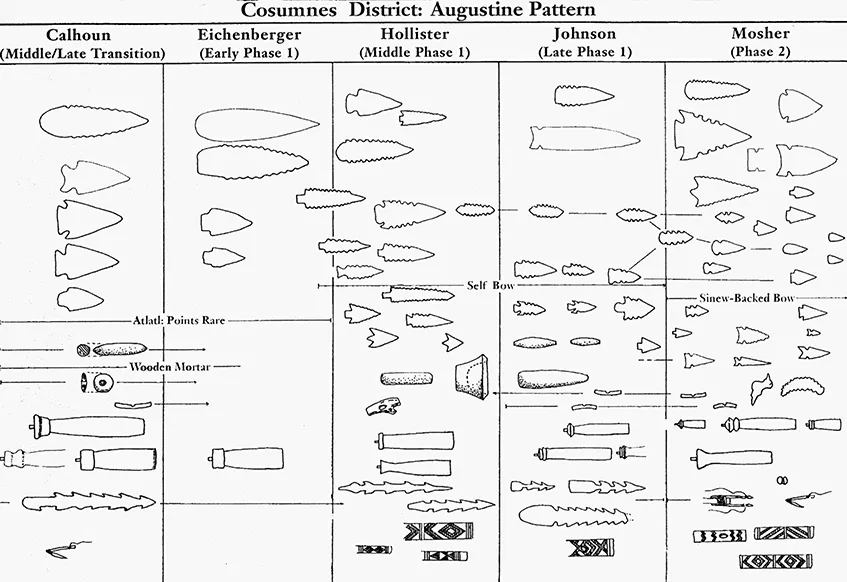
Another example is James Bennyhoff’s (1994) map of the relations of Central Valley, California, projectile points, beads, and amulets (figures 1.3 and 1.4). Bennyhoff detailed change in these artifact classes through time and linked cases where he believed divergence occurred among artifacts (as with the amulets and beads) as well as convergence (as with the projectile points). These time-space charts are an embedded feature of archaeology and provide a framework for studying culture change. And for good reason: whenever our goals include explaining change through time, it is necessary to build models of relatedness. These models isolate characters suspected to be the result of inheritance as they vary through time and across space. requisite when testing hypotheses about the role of environments in structuring variability in cultural behavior. As the zoologist Paul Harvey (1996: 257) has noted, “if we want to understand why different traits are more commonly represented in one community than another we shall frequently find phyloge-netic information useful. It can only help, and it will produce statistically appropriate degrees of freedom.”
Figure 1.2
Development of Manos and Metates at Snaketown, Arizona (after Sayles 1937)
Accurately determining the degree to which the sharing of traits is a function of historical relatedness is vital in building evolutionary explanations. In order to explain the distribution of cultural traits across populations we must be able to identify those traits that are present as a result of historical contingency (“homologies”) versus those that are a product of processes other than descent (“homoplasies”). Consider a case of explaining why projectile points found distributed in densely wooded upland environments are different from those found along oak-savannah valley bottoms.2 We might find, for example, that some of the points in the uplands are slightly smaller than contemporaneous ones in the lowlands. A chi-square test might demonstrate statistical significance in the pattern, and we might therefore be tempted to explain the differences as a function of the different environments. Such an explanation is plausible but potentially wrong. It is possible that the lowland and highland groups may have inherited their preferences regarding projectile-point length from different common ancestors. If this were the case, it would be inappropriate to treat the groups as independent data points in a statistical analysis (Harvey and Pagel 1991). Reconstructing phylogeny is therefore a vital pre-
Figure 1.3
Historical Evolution of Beads and Amulets in the Central Valley, California (after Bennyhoff 1994)
Figure 1.4
Historical Evolution of Projectile Points in the Central Valley, California (after Bennyhoff 1994)
Culture-historical time-space charts are definitely the sort of product we need. However, the means of generating them must be vastly improved. We must be able to build maps of relatedness in ways that are theoretically justified, reproducible, and quantitatively defensible. The majority of culture-historical depictions of patterns of descent are little more than intuitive claims about relationships based on experience and authority. Although these maps do a reasonable job of determining large-scale differences (for example, distinguishing between early and late manifestations), they are incapable of resolving small-scale differences. At best, they are nominal-level representations of relatedness. Here is a significant reason for us to care about phylogenetics: phylogenetic methods such as cladistics offer us a means of systematically deriving theoretically justifiable maps of relatedness using explicit algorithms in a way that is repeatable. These methods are integral to all forms of evolutionary explanations, whether biological or cultural, since the central tenet of evolution, descent with modification, requires us to track related entities as they change through time and across space. Without showing relatedness, explanations of change are simply chronological, not evolutionary, statements. Thus, phylogenetic methods are central to our quest for explaining the natural world.
Issues in Cultural Phylogenetics
Given the requirements of evolutionary studies, regardless of whether the subject matter is language, artifacts, or social institutions, we see phylogenetics as an important component. Critics, however, have not only raised a number of issues with respect to the appropriateness of various methods for unraveling cultural phylogenies but have also questioned whether cultural phylogenies can even be understood, regardless of the method used. If the criticisms are valid, they not only limit the applicability and effectiveness of various phylo-genetic methods, they call into question the entire phylogenetic enterprise. We summarize some of these issues below, leaving it to various chapter authors to address them in more detail.
Culture versus Genes
Some researchers argue that phylogenetic methods are inappropriate for studying cultural evolution because they rest on a false analogy. This argument is not new (e.g., Brew 1946). It holds that cultural transmission is not analogous to genetic transmission—that unlike with genes, culture is not a transmission system in which physically identifiable entities are passed from person to person structurally intact (Atran 2001; Aunger 2000; Sperber 1996). As such, we cannot directly apply biological methods to cultural data. In an intuitive sense this argument appears defensible. Obviously the “things” that pass between people in a cultural-transmission system are not sharply defined objects. However, simply because we cannot see the transmitted “things” does not mean that we cannot see their effects.
Sometimes cultural transmission is said to take place via gene-like units called “memes” (Dawkins 1976). Although the concept of meme enables us to conceptualize a unit for measuring cultural transmission, we agree with Lake (1996) that much of the memetics literature creates confusion by conflating the physical expression with the content being transmitted (e.g., Blackmore 1999; Gabora 1996). The example of an image or string of words, copied and passed around while retaining its essential identity, is commonly used as a memetic example of cultural transmission. This image is inappropriate.
If physical expression is not part of the process of cultural transmission, then what does transmission consist of? What is passed on? In a word, information is passed on. Clearly, we are not the first to say this (e.g., Cloak 1973, 1975; Dawkins 1976). But in order to address the criticism that cultural evolution is at best analogous to genetic evolution (and then only vaguely so), we have to be clear that transmission is about the passage of information between individuals at whatever scale and using whatever physical means is available (chemical, molecular, sound, or light). Thus, there cannot be a single, physical entity in any system of information transmission. There are no “strings of words.”
It is important to recognize that the lack of a single physical entity is true even in the case of genetic transmission, where there are numerous physical entities—DNA, transfer RNA, and many proteins—that carry and pass on information. As we have delved deeper into the mechanisms of genetic transmission, we have learned that DNA does not play an exclusive role in transmitting information between individuals. Similarly, in cultural transmission there are also a number of physical mechanisms that result in the transmission of information.
Even though we might not all agree on the mechanism or even on what is transmitt...