
- 394 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Originally published in 1987 this title was designed as a textbook for first degree students of psychology and provides an introduction to the major topics within the subject of physiological psychology. The aim was to cover these major subject areas and at the same time to provide indications of advances made in the previous two decades. Today the book is still suitable for all levels of study, from beginning students to final year level, who wish to cover historical aspects of physiological psychology.
Tools to learn more effectively

Saving Books

Keyword Search

Annotating Text

Listen to it instead
Information
1 The nervous system – basic principles of function and techniques of investigation
Physiological psychology is concerned with the links between the organism’s physiology and its behaviour. Physiology incorporates the totality of physiological systems within the body – the nervous system, endocrine system (glands secreting within the body, such as thyroid, pituitary, adrenal glands), exocrine system (glands secreting via tubes, or ducts, such as sweat glands and tear ducts), skeletal muscles, the gastrointestinal system, the circulatory system, etc. Although we shall be meeting some of these systems in later chapters, the predominant interest of the physiological psychologist lies in the brain and the rest of the nervous system. The basic unit of the nervous system is the neuron.
The neuron
All the cells of the body follow the same basic pattern, with a nucleus containing chromosomes, the genetic material consisting of strands of deoxyribosenucleic acid (DNA), which is surrounded by the jelly-like cytoplasm containing all the other cellular mechanisms, e.g. the mitochondria, for energy production, and the ribosomes, for protein synthesis. Every cell in the body contains an identical set of chromosomes. Besides embodying an organism’s characteristics and controlling their transmission from generation to generation, the chromosomes and their DNA control the cell containing them. In some as yet unexplained fashion, they enable different cells to specialize in different ways during development, despite having the same basic pattern. The muscle cells become specialized for contraction, gland cells for secretion, and neurons for the transmission of electrical impulses.
The particular functional specialization of the neuron leads to physical specializations which can be obvious or very subtle. Neurons are elongated (see Fig. 1.1), with the cell body (‘soma’) extending into a relatively long branching axon on one side and into a number of relatively short dendrites on the other. It is important to remember that the whole neuron is a single cell, with cytoplasm filling axons and dendrites.
Neurons come in a wide range of sizes and show an infinite variety of dendritic and axonal processes. The human brain contains of the order of fifteen billion neurons (15 × 109), most of which will have short dendrites and axons and be invisible to the naked eye. On the other hand, the axon transmitting electrical impulses from the spinal cord to the muscles of the toes belongs to a neuron which can be three to four feet long, depending on the length of your legs.
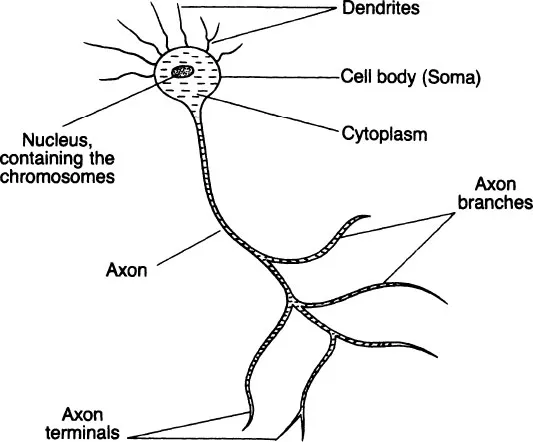
Figure 1.1 The neuron
The subtle specialization all neurons have in common concerns the cell membrane, the outer barrier of the neuron. This is semi-permeable, allowing the passage of various electrically charged particles (ions) from inside to out and from outside to in. Usually there is a differential concentration of ions inside and out, leading to an electrical difference, or potential, of around – 70 millivolts across the membrane from inside to out. The situation can be altered by stimuli of various sorts, especially small electric currents. Any stimulus which shifts the membrane potential towards equilibrium (i.e. zero membrane potential) may provoke an action potential at that point on the membrane. As the potential across the membrane is decreased to around –50 millivolts, there is an explosive increase in membrane permeability, enabling sodium and chloride ions to rush into the neuron and potassium ions to pass out; this rapid exchange shifts the membrane potential from -50 to around +40 millivolts in about half a millisecond, and constitutes the action potential.
After the action potential has peaked at +40 millivolts, the membrane rapidly returns to its resting state of a –70 millivolt potential. With a brief refractory period after the action potential during which the membrane is inactive, the whole sequence takes about four milliseconds, i.e. the temporal gap between action potentials must be at least four-thousandths of a second.
This gap becomes important when we consider what happens to the action potential. The structure of the neuronal membrane that enables action potentials to occur also enables them to travel along the neuron from the point at which they were first stimulated. Thus a wave of electrical activity (or ‘depolarization’, the explosive change in membrane potential from –70 to +40 millivolts) passes along the neuronal membrane. If a thin wire recording electrode is placed on the membrane, it will record this wave as a blip of electrical activity; this is the nerve impulse. If action potentials are successively stimulated, they will be recorded as sequences, or trains, of nerve impulses. Each part of the membrane traversed by an impulse will be inactive for around four milliseconds. Thus the maximum rate of impulse transmission, or frequency, is around 250/second. As one action potential is electrically identical to any other, each impulse is similarly indistinguishable from every other impulse. The crucial properties of impulse conduction are therefore the frequency and temporal patterning of trains of impulses.
Action potentials are usually initiated on the dendrites of the neuron and the nerve impulse is then propagated along the neuron in the direction dendrite → soma → axon → terminals of all axon branches. Before considering the initiation of the action potential in more detail, there is one fundamental point to be made. Information in any nervous system is embodied, or coded, entirely by the frequency and patterning of nerve impulses. Sensory and motor systems, perception, memory, cognition and thought, emotion, personality, etc., are all represented by patterns of nerve impulses in appropriate parts of the nervous system.
This can be simply demonstrated. Using thin wire stimulating electrodes, a tiny electric current can be applied to single neurons or groups of neurons in the brain. As the brain itself has no sensory receptors, this procedure can be performed in human patients using only a local anaesthetic for the area of the head and skull through which a hole will be drilled to allow entry of the electrode into the brain. Such techniques are normally part of a screening programme before major brain surgery.
If the stimulating currents are within the naturally occurring limits for nervous system electrical activity, then the normal function of an area of the brain can be mimicked. As the patient is conscious, he or she can report any subjective experiences. Over the years it has been shown that visual experience (flashes of light), auditory sensation, ‘automatic’ vocalization, feelings of unease and euphoria and outbursts of aggression, recall of memories, movement of limbs and of individual muscle groups, increases and decreases in heart-rate and blood pressure, can all be elicited by electrical stimulation of the appropriate part of the brain.
So we possess a brain of unimaginable complexity using a fundamentally simple electrical code for the information represented within it. Every aspect of behaviour and physiology discussed in this volume has its essence in patterns of identical action potentials in the nervous system, i.e. if you want to be reductionist about it, our brains can be represented as machine-like. However, this machine has fifteen billion working units each, as we shall see, having several thousand interconnections with other units, and produces human behaviour and experience; it is only machine-like if we upgrade our concept of what machines are and what they can do, as even the most sophisticated computer can do little, apart from sums, that we would recognize as a truly human faculty.
The synapse
Neurons are not physically connected to one another. Otherwise, action potentials would simply travel haphazardly to all parts of the nervous system. Between an axon terminal and the next neuron is a gap, the synapse (see Fig. 1.2). Only visible under the electron microscope, the synapses are measured in angstroms (an angstrom is one-millionth of a metre), and present a barrier to the action potential travelling down the axon to the axon terminal (or pre-synaptic terminal); the action potential must be transmitted across the synapse to the postsynaptic membrane of the next neuron if it is not to be lost.
Transmission across the synapse is chemical. Quantities of the particular chemical (‘synaptic neurotransmitter’) are stored within the pre-synaptic terminal in spherical storage vesicles. When an action potential reaches the pre-synaptic region, it stimulates a number of vesicles to migrate to and merge with the actual neuronal cell membrane bordering the synapse, where they release their chemical contents into the synaptic cleft. The molecules of neurotransmitter diffuse across the synapse and combine with ‘receptors’ located on the postsynaptic membrane. This combination is most simply seen as a ‘lock-and-key’ affair. The molecule of transmitter has a certain spatial configuration which is assumed to match perfectly the spatial configuration of the receptor molecule.
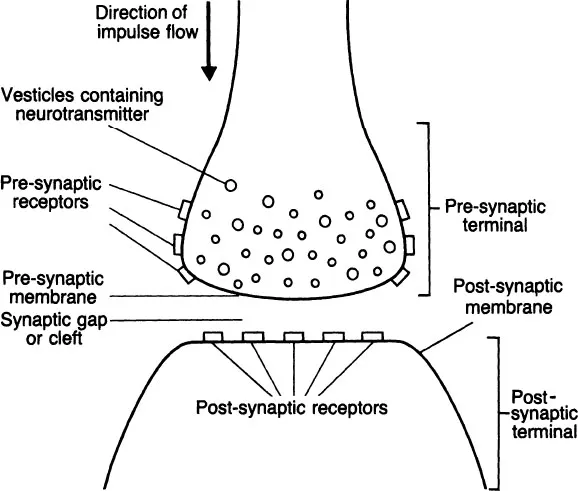
Figure 1.2 The synapse
This interaction between transmitter and receptor is shortlived, but while it lasts produces a change in the permeability of the post-synaptic membrane. This change is along the lines described for the initiation and propagation of the action potential, but the combination of a single molecule of neurotransmitter with a single receptor is insufficient to fully depolarize the post-synaptic membrane. An action potential is based upon depolarization, and even the total amount of neurotransmitter released by a single action potential in the pre-synaptic terminal will not be sufficient to initiate an action potential in the post-synaptic neuron, i.e. the nerve impulse will not cross the synapse, and the information it represents will be lost.
For synaptic transmission to occur, sufficient nerve impulses must arrive within a short space of time at the pre-synaptic terminal to stimulate the release of a sufficient quantity of neurotransmitter into the synapse; when combining simultaneously with many post-synaptic receptors, this quantity will be enough to depolarize the post-synaptic membrane and so initiate an action potential which will in turn be transmitted along the post-synaptic neuron.
Thus the frequency of nerve impulses in the pre-synaptic neuron is unlikely to be matched by the frequency in the post-synaptic neuron. There is one obvious function for this mechanism, but also a further complication. The obvious function is the filtering out of irrelevant information. If touch receptors on the skin are momentarily activated by dust particles in the air, nerve impulses travel along sensory neurons towards the central nervous system. If the activation is momentary, the concentration of impulses will be insufficient to carry the first synapse in the chain leading to the brain and conscious awareness. Thus the information will be lost, or filtered out, a quite reasonable system for preventing the brain becoming overloaded with what is fundamentally irrelevant information.
The complication in the mechanism is that synaptic connections are not one-to-one, or even of the same type. As a very rough estimate, any central nervous system neuron makes contact via synapses with several thousand other neurons. Thus a nerve impulse in one neuron is the summated product of hundreds or thousands of inputs to that neuron; the nervous system does not operate along straight lines.
Neurotransmitters
Chapter 3 considers brain chemistry and behaviour in some detail. At this stage I want to make some general points regarding neurotransmitters in the brain.
Chemical neurotransmission was first demonstrated by Loewi in 1921 (following, incidentally, ‘creative’ dreaming!) using the neural connections to the frog’s heart (Loewi and Navratil, 1926). Over the next thirty years work concentrated upon the peripheral nervous system, particularly the synapses between the axons of motor neurons and skeletal muscles (the neuro-muscular junction), and it is still the case today that our synaptic models are based upon the peripheral nervous system or the accessible nervous systems of other animals. Technical problems rendered the central nervous system (brain and spinal cord) more or less impervious to pharmacological investigation, and as late as 1950 it was still quite acceptable to deny the existence of synapses in the brain, i.e. to believe that transmission was entirely via electrical conduction along and between neurons.
Chemical neurotransmission at a central nervous system synapse was first demonstrated in the spinal cord (Eccles et al., 1954), and rapidly became accepted as a fundamental means of information transmission in the brain. (By ‘demonstrated’ I mean that certain criteria were established, e.g. presence of neurotransmitter in pre-synaptic terminals, its release in response to nerve impulses, presence of post-synaptic receptors with which the neurotransmitter can combine, relevant electrophysiological changes, etc.).
By 1970 there were six established chemical neurotransmitters: noradrenaline, dopamine, and serotonin (all monoamines), acetylcholine (tertiary amine), GABA (gamma-amino butyric acid) and glutamate (both amino acids). It is a basic principle, now under attack but still not fully disproved, that a given neuron will secrete the same neurotransmitter at all of its axonal terminals (Dale’s principle), and that neurons may therefore be characterized by their neurotransmitter. Thus the brain contains noradrenergic, dopaminergic, serotonergic, cholinergic, GABA-ergic, and glutamatergic neurons.
Added to this list by now are anywhere between twenty and thirty possible neurotransmitters discovered over the last decade, some of which are discussed in Chapter 3 and whose precise role in brain function has not yet been identified. Some may be synaptic neurotransmitters in the classic sense, while others may be ‘neuromodulators’, influencing neurotransmission in subtle ways. Regardless of the ultimate outcome, brain chemistry has become the fastest-growing area of brain re search, partly because the existence of a chemical bridge between neurons provides a valuable means for influencing brain function using externally applied chemicals and for modelling drug effects on behaviour, and partly through a combination of sophisticated biochemical and pharmacological techniques which have only become available in the last fifteen years or so.
I mentioned earlier that synaptic receptors were specific to a given neurotransmitter. So, in line with the different transmitters there are dopamine, noradrenaline, acetylcholine, etc., receptors. I also pointed out that neurons receive many inputs via synapses, and so their dendritic and somatic membranes may possess many different types of receptor depending upon the types of input.
Two further complications. An axon terminal releases neurotransmitter into the synaptic cleft where it diffuses over to the post-synaptic membrane and combines with its specific receptors. It has been discovered that there are also receptors on the pre-synaptic terminal itself; some of the neurotransmitter diffuses to these receptors (‘autoreceptors’), where the effect is to inhibit neurotransmitter release from the pre-synaptic terminal, i.e. a negative feedback system, as the more transmitter is released the less likely it is to be released.
Secondly, neurotransmitter receptors exist in several forms. There are muscarinic and nicotinic cholinergic receptors, alpha and beta noradrenergic receptors, Dl, D2 and D3 dopaminergic receptors (with several more awaiting confirmation), and serotonin 1 and 2 receptors. The subtypes all combine with the neurotransmitter itself, but are distinguished by their ability to combine with (or bind to) other drugs. So the muscarinic cholinergic receptor is stimulated by acetylcholine and the drug muscarine, but not by nicotine, while the nicotinic cholinergic receptor is stimulated by acetylcholine and nicotine, but not by muscarine.
Receptor subtypes may also be distinguished by their synaptic or regional distribution. Thus noradrenergic pre-synaptic autoreceptors are of the alpha type, while all synapses at the skeletal neuromuscular junction are of the nicotinic cholinergic type. This amazing chemical heterogeneity makes the inter pretation of drug effects extremely difficult, and while I have mentioned it here to illustrate the rapid pace of brain research, I do not intend to go into it more deeply than is necessary in subsequent chapters. There are some fine introductory texts (e.g. Iversen and Iversen, 1981) which cover the chemistry and pharmacology of brain function in impressive detail.
Techniques for investigating brain function
The two principles of information transmission in the nervous system – electrical and chemical – immediately provide two sets of investigatory techniques.
Electrical stimulation and recording
I have already mentioned the use of electrical stimulation to mimic the brain’s natural activity. This necessarily involves localized stimu...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Original Copyright Page
- Dedication
- Table of Contents
- Acknowledgments
- Preface
- 1 The nervous system – basic principles of function and techniques of investigation
- 2 The nervous system – organization and functions
- 3 Brain chemistry
- 4 Sensory systems
- 5 Functions and asymmetries of the cerebral hemispheres
- 6 Brain mechanisms of memory and learning
- 7 Habituation – attention and memory
- 8 Emotion – physiological psychology and central mechanisms
- 9 Emotion – psychophysiology and peripheral arousal systems
- 10 Central mechanisms of arousal, sleep, and waking
- 11 Motivation
- 12 Brain chemistry and psychopathology
- References
- Subject index
- Selected author index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Physiological Psychology by Simon Green in PDF and/or ePUB format, as well as other popular books in Psychology & History & Theory in Psychology. We have over one million books available in our catalogue for you to explore.