This volume contains a collection of papers written by former students, postdoctoral fellows, and colleagues of Richard Thompson and represent written versions of papers presented at the Festschrift symposium. The Festschrift provided an excellent opportunity for the participants to recount their memories and experiences of working with one of the leading figures in behavioral neuroscience, and to place their current research in the context of earlier research conducted in the Thompson laboratory. As a Festschrift volume, the various chapters contain numerous and sometimes very personal references to Richard Thompson's influence on the careers of the authors, as well as summaries of past and present work being conducted in the authors' laboratories. Part I includes studies of spinal cord plasticity and the involvement of the hippocampus and related structure in classical eyeblink conditioning. Part II explores the critical role of the cerebellum and associated areas in classical eyeblink conditioning. Part III focuses on a continued exploration of the involvement of the cerebellum in classical eyeblink conditioning using standard procedures as well as innovative molecular biology and genetic techniques. It also includes studies aimed at delineating modulatory influences on learning such as stress and hormonal factors. The incredible influence that Richard Thompson has had on the fields of experimental psychology and neuroscience should be evident on reading the contributions made by the various authors to this volume. The research conducted in Thompson's laboratory over the years has been cutting-edge, comprehensive, and influential. Therefore, this volume is dedicated to Richard F. Thompson a productive, innovative scientist and outstanding mentor.

eBook - ePub
Model Systems and the Neurobiology of Associative Learning
A Festschrift in Honor of Richard F. Thompson
- 440 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Model Systems and the Neurobiology of Associative Learning
A Festschrift in Honor of Richard F. Thompson
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
1
Classical Conditioning of Spinal Reflexes: The First Seventy Years
The neural makeup and basic functions of the mammalian spinal cord have been well recognized for many years. Early in the last century, Sherrington (1905) studied spinal reflexes and recognized that they could undergo response increases or decrements (reflex fatigue) with repeated use. However, the common view for the first part of the 1900s was that the mammalian spinal cord was essentially a passive transmitter of information and that information processing of any import was confined to the brainstem and cortex. This viewpoint constrained most research into spinal function to investigating and cataloging reflex functions of the spinal cord. The first epoch of spinal cord classical conditioning began in the late 1930s when a few investigators began to examine spinal reflexes in response to the question "What is the most elementary neural circuits of the nervous system that can support learning." The reflex circuits of the cord were readily accessible for stimulus delivery and response recording, and could be isolated from the brainstem by spinal transection. Early studies struggled with the questions, not yet fully resolved, of a definitive definition of learning and whether a behavioral process acceptable to learning theorists could be found in the spinal cord. The dialogue beginning in the late 1930s which continues to some extent today began to produce evidence that spinal neural circuitry was more plastic and capable of more change than previously thought.
In the mid 1960s, Thompson and his colleagues (Glanzman, Teyler, & Thompson, 1975; Thompson, Patterson, & Teyler, 1972; Thompson & Spencer, 1966) began to formalize the concepts of utilizing simplified systems as models for investigating learning in intact animals or humans. This led to the second epoch of spinal classical conditioning studies, from the mid 1960s through the late 1980s. The ideas of "model systems" and "model learning phenomena" were conceptually very useful in furthering the search for both behavioral and neural substrates of behavioral change. These ideas and the information gathered from the studies of spinal reflexes as model systems led to the development of the third epoch in spinal cord conditioning research that began in the late 1980s and is now in full swing. With the realization that spinal reflex systems were not simple, hard-wired, unchanging entities, the picture now emerging is one of spinal neural circuits actively engaged in shaping behavior and even sensory inputs (Patterson, 1980b). As increasing amounts of information are gathered about spinal cord function, it is becoming clearer that a variety of functions ranging from locomotor activities to pain perception and visceral function are affected by active processing of information at the spinal level. Thus, the current view of spinal cord function is vastly different now than it was even 25 years ago.
The present chapter examines the use of classical conditioning techniques to investigate the neural potentials of spinal cord function. The studies of "spinal conditioning" have mirrored the epochs outlined earlier and have contributed to the changing notions of spinal cord capabilities. The implications of understanding spinal function are vast, ranging from a fuller understanding of normal motor control, through better means of pain control, to vastly improved rehabilitation of spinal cord injuries.
CLASSICAL CONDITIONING
The basic concepts of classical conditioning were introduced by Pavlov (1927). The concepts developed over the early part of the century by this Russian physiologist flowed from a realization that by studying behavioral changes in response to environmental stimuli, inferences about underlying brain function could be made. Pavlov developed the concept of an unconditioned stimulus (UCS), a stimulus that was prepotent or inherently capable of causing a response in the target organism. Thus, the UCS of mild acid delivered to the oral cavity would, without prior experience, produce salivation (the unconditioned response, or UCR) in an animal. The signal stimulus, or conditioned stimulus (CS) on the other hand, would not initially produce a response in at least the target response system. By associating the two stimuli in time, a response would often come to be elicited by the CS; the conditioned response or CR. Pavlov viewed the CR to be the result of neural alterations at the cortical level due to the association of the CS with the prepotent UCS. He viewed this associative process as the basic learning mechanism of mammals and with it developed empirical and theoretical explanations for human and animal behaviors ranging from simple learning to various mental disorders.
American psychologists of the 1930s began to investigate classical conditioning as a means to understand learning. The apparently simple phenomenon of the association of a CS with a UCS seemed to be about as simple as a learning process could be. This simplicity is, however, somewhat deceptive, but nevertheless, soon led to the question of how much neural organization was necessary in the mammalian system to support this simple learning. An obvious place to look for the answer was in the reflex systems of the mammalian spinal cord.
Classical Conditioning of Spinal Reflexes: The First Epoch
The initial studies of spinal conditioning have been extensively reviewed (Patterson, 1976). I briefly review the major ones here.
Pavlov viewed the great cerebral cortices of the mammal as the seat of learning. Elmer Culler and his colleagues at the University of Illinois became interested in the question of whether a mammal deprived of its cortex could show behavioral changes that could be called learned. Poltyrev and Zeliony (1929) had reported that decorticate dogs could show behavioral changes consistent with learning. Culler (Culler & Mettler, 1934; Girden, Mettler, Finch, & Culler, 1936) produced evidence that decorticate dogs could show conditioned responses. However, the responses lacked the fine tuning associated with normal conditioning of motor responses. These studies led directly to the first attempts to produce spinal conditioning (Culler, 1937). Young dogs were spinalized and a CS of pressure or electric shock to the tail tip was paired with hindpaw shock as the UCS. The result seemed to be the development of contractions to the CS in the measured semidendinosis muscle of the shocked leg (for a full report, see Shurrager & Culler, 1940). In this report they set the stage for the controversy that followed. They stated that the data showed that the cord was capable of acquiring substitute responses that were not reflexively present initially in the isolated cord. The implication was that, as was believed to be the case in the cortex with classical conditioning procedures, the delivery of CS and UCS to the spinal reflex pathways had created a "new" reflex. Fig. 1.1 shows data from 57 of the over 200 dogs conditioned. The delivery of the CS-alone following the training trials in the classical conditioning paradigm generally results in the decrease of the CR called extinction, as seen in the figure. In a later paper (Shurrager & Shurrager, 1946), evidence was presented that repeated conditioning, extinction, reconditioning sessions increased the speed of reacquisition of the CR. The data were taken to support the notion that the mammalian spinal cord could support at least the simple associative learning of classical conditioning.
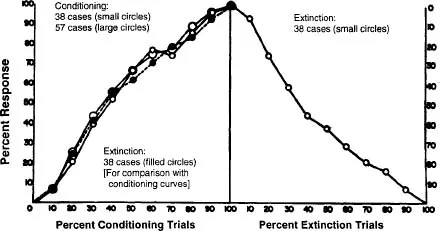
FIG. 1.1. Vincent-curves of original conditioning and extinction in the spinal dog. Small open circles, conditioning, and subsequent extinction in 38 preparations. Large open circles, conditioning in 57 preparations. Solid circles and dotted line, extinction curve turned vertically through 180° and superimposed upon the conditioning curves for comparison. From "Conditioning in the spinal dog" by P. S. Shurrager and E. Culler, 1940, Journal of Experimental Psychology, 26, p. 136. Copyright 1940 by the American Psychological Association, Reprinted with permission.
Culler's interpretations were soon challenged by Kellogg and his colleagues (Kellogg, Pronko, & Deese, 1946; Pronko & Kellogg, 1942). In these studies, Kellogg found what he interpreted not as associative learning in the spinal animal, but rather simple modification of existing reflexes. On this argument turned the question of spinal conditioning. While Kellogg and his associates found, at least in some instances, alterations of a response to the CS, they took the view that this was not the production of a "new response pattern" as required by Pavlovian dogma.
The conflicts between the two early groups of investigators were apparently due to both theoretical and practical considerations. The methods used by the two groups were not the same. In many cases, the Kellogg group used chronic preparations, whereas the Shurrager group used acute preparations. The Kellogg group used a shock CS to one hind foot and the shock UCS to the other. This could have produced competing response tendencies during conditioning to the CS. These and other problems (see Patterson, 1976, for a full discussion) left the question of spinal conditioning open at the end of the initial epoch of spinal conditioning research.
Refinement of Classical Conditioning
The Pavlovian paradigm of classical conditioning, as with any experimental paradigm, underwent a continual refinement. As initially practiced, classical conditioning was simply the observation of response development with the pairing of the CS and UCS, and decrement of the resulting CR to repeated CS-alone (extinction) presentations. Although seemingly simple, the experimental procedures of classical conditioning require close attention. One of the problems evident in the early spinal conditioning studies was that of determining whether the actual temporal pairing of the two stimuli was the crucial factor in any observed behavioral change. In 1962, Gormezano and his colleagues (Gormezano, Schneiderman, Deaux, & Fuentes, 1962; Schneiderman, Fuentes, & Gormezano, 1962) introduced what has become one of the most widely utilized animal preparations for the study of behaviors and neural substrates of classical conditioning, the rabbit nictitating membrane model. Gormezano also introduced the most universal control condition for classical conditioning: the unpaired control. Reasoning that the simple presentation of two stimuli could alter response patterns, Gormezano argued that by contrasting the behavior of subjects given the CS and UCS temporally paired with animals given the same stimuli presented the same number of times but never together in time, a true measure of learning could be obtained. Although oversimplified, this argument was the basis for a revolution in modern classical conditioning procedures. The use of an explicitly unpaired control group allowed a much more precise determination of the effects of classical conditioning procedures than previously possible. Further refinements in the planned control group procedures have been made, such as the implicitly unpaired control group, Here, the two stimuli are allowed to occur at random, with the result that an occasional paired trial is possible. However, the introduction of the unpaired control allowed greatly increased precision in determining the extent of learning due to classical conditioning.
Classical Conditioning of Spinal Reflexes: The Second Epoch
The introduction of planned control group technology into classical conditioning quickly led to the first spinal conditioning study utilizing this control. Fitzgerald and Thompson (1967) reported a study in which they utilized decerebrate, spinalized cats given a shock CS to the left thigh skin paired with a shock UCS to the left toe pads. The cats given paired trials showed marked increases in the leg flexion response to the CS while the cats given unpaired CS and UCS presentations showed no such response increases. The CRs of the paired group decreased with extinction trials. Fig. 1.2 shows the results of the paired experimental and unpaired control group responses during acquisition and extinction trials. This study also introduced the use of a CS that initially and intentionally produced a small response prior the conditioning. This alteration in procedure acknowledged the fact that in the spinal reflex system, the use of a CS that produced no response output could not reliably be determined to be received by the spinal circuits. This change acknowledged that the behavioral alteration of interest was the effect of stimulus pairing on a pre-existing response, not the creation of a new response.
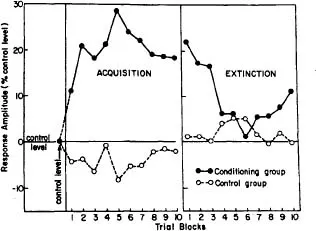
FIG 1.2. Performance of the conditioning and control groups during acquisition and extinction plotted as percent change from initial control level. Redrawn from "Classical conditioning of the hindlimb flexion reflex in the acute spinal cat" by L. A. Fitzgerald and R. F. Thompson, 1967, Psychonomic Science, 8, p. 8. Copyright 1967 by the Psychonomic Society. Reprinted with permission.
The confluence of the new insights into classical conditioning procedures and the alteration of the spinal conditioning paradigm to examine not the creation of a new response but the specific effects of pairing on an existing response began a renewed interest in spinal conditioning. In 1968, Buerger and Dawson (1968) reported the results of a study of chronic spinal kittens (2-15 weeks post spinal section) given a tail pinch CS and toe pinch UCS. The results of paired versus unpaired stimulus groups showed an EMG increases over training sessions in the paired group with only response habituation in the unpaired group. This was significant in that the subjects were midterm chronic animals, as opposed to the acute preparations most previous studies (except the Kellogg studies) had used.
Thompson, in designing his study of spinal classical conditioning, realized that the preparation had potential use not only in determining the ability of spinal circuits to support learning or learning-like behavioral alterations, but also as a means to investigate the neural substrates of these behaviors. He and his colleagues had begun studies of reflex habituation and sensitization in the spinal preparation some time before (Spencer, Thompson, & Neilson, 1966; Thompson & Spencer, 1966). That work showed that spinal reflexes were capable of many attributes of similar behaviors in traditional experimental psychology. In addition, the spinal cord was considered to be much more amenable to analysis of neural mechanisms than the intact mammalian brain.
The Preparation
In 1969, we began our collaboration with Thompson in further developing the spinal classical conditioning preparation and paradigm. The spinal preparation was refined to provide maximum utility for both behavioral and neural mechanism investigations. Our pilot studies determined that the preparation could utilize a cat spinalized under ether anesthesia, then paralyzed with Flaxedil, artificially respirated and ether d...
Table of contents
- Front Cover
- Half Title
- Title Page
- Copyright
- Contents
- List of Contributors
- Preface: The Career and Scientific Contributions ix of Richard F. Thompson
- 1 Classical Conditioning of Spinal Reflexes: The First Seventy Years
- 2 Forms of Associative Synaptic Plasticity
- 3 The Ins and Outs of Classical Conditioning: Maxims From a Mollusk
- 4 The Septo-Hippocampal System and Classical Conditioning
- 5 Model Systems and Memory: Applications and Extensions to Clinical Neuroscience
- 6 A Tale of Two Paradigms: Lessons Learned From Parallel Studies of Discriminative Instrumental Learning and Classical Eyeblink Conditioning
- 7 Receptive Field Plasticity and Memory in the Auditory Cortex: Coding the Learned Importance of Events
- 8 The Cerebellum and Classical Eyeblink Conditioning: Much Ado About Something
- 9 Inside the Black Box
- 10 The Cerebellum and Associative Learning: Parallels and Contrasts in Rabbits and Humans
- 11 Medial Septal Modulation of Conditioning: From Two-Stage Theories to Connectionist Models
- 12 Estrogen and Synaptic Plasticity in the Hippocampus
- 13 The Modulation of Memory Formation by Stressful Experience and Sex Differences in the Brain
- 14 Eyeblink Conditioning Impairments in Patients with Motor Disorders
- 15 Toward a Neurobiology of Functional Behavior Systems: Contrasting Pavlovian Emotional and Motor Learning
- Author Index
- Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Model Systems and the Neurobiology of Associative Learning by Joseph E. Steinmetz, Mark A. Gluck, Paul R. Solomon, Joseph E. Steinmetz,Mark A. Gluck,Paul R. Solomon in PDF and/or ePUB format, as well as other popular books in Psicología & Neurociencia cognitiva y neuropsicología. We have over 1.5 million books available in our catalogue for you to explore.