This book examines how the growing knowledge of the huge range of protist-, animal-, and plant-bacterial interactions, whether in shared ecosystems or intimate symbioses, is fundamentally altering our understanding of biology. The establishment and maintenance of these interactions and their contributions to the health and survival of all partners relies on continuous cell-to-cell communication between them. This dialogue may be concerned with all aspects of the biology of both partners. The book includes chapters devoted to exploring, explaining, and exposing these dialogues across a broad spectrum of plant and animal eukaryotes to a broad field of biologists.
Key Features:
Explores the nature of the interactions between eukaryotic hosts and their microbial symbionts
Examines the links between prostist, animal, and plant evolution and microbial communities
Reviews specific taxa and the microbial diversity associated with these taxa
Illustrates the role microbes play in the physiology and etiology of several model species
Includes chapters by an international team of leading scholars
The Open Access version of this book, available at http://www.taylorfrancis.com, has been made available under a Creative Commons Attribution-Non Commercial-No Derivatives (CC-BY-NC-ND) 4.0 license.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
When does symbiosis begin? Bacterial cues necessary for metamorphosis in the marine polychaete Hydroides elegans
Marnie Freckelton and Brian T. Nedved
Contents
1.1 The symbiosis space
1.2 Chemical cues mediate symbiotic interactions
1.3 How do specific symbiotic interactions begin? Examples from the pre-symbiosis space
1.4Bacterially induced metamorphosis of marine invertebrate animals
1.5 Bacterial induction of metamorphosis in Hydroides elegans
1.6 Identification of larval metamorphic cues from biofilm bacteria
1.7 How variability of inductive bacteria and identified settlement cues relate to variable larval settlement and recruitment
1.8 Lipopolysaccharide mediates both symbiotic and pre-symbiotic interactions
1.9 Conclusion
References
1.1 The symbiosis space
Symbiosis, the living together of unlike organisms (de Bary 1879), has become one of the most important concepts in biology today. This has been especially true since the discovery of the prevalence of microbiomes (Woese 2004; Moran and Dunbar 2006) and the role of the holobiont (Margulis 1971) in shaping both individual health and development, the evolution of biological complexity, and ecosystem-wide processes (Bordenstein and Theis 2015). It is through this cooperation of individuals that new habitats and energy sources can be accessed and utilized (Moran and Dunbar 2006; Fisher et al. 2017). In the marine environment, cooperative prokaryotic-eukaryotic interactions have drastically increased the rate at which organisms adapt to utilize new environments and energy sources (Apprill 2020).
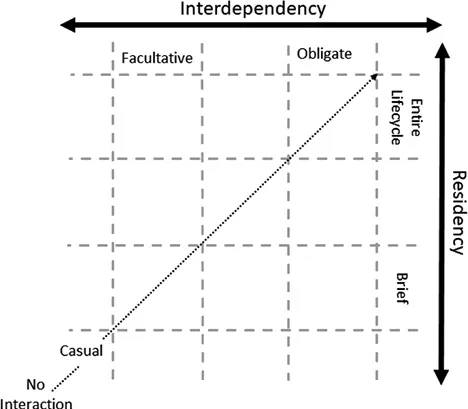
Symbiosis, however, is not a discrete concept but rather extends across a spectrum of interactions defined by three axes: impact on host fitness (positive to negative), the dependency of the relationship, and the residency of the symbiotic relationship. In prokaryotic–eukaryotic symbioses, the evolutionary relationship is considered to start with complex, nonessential interactions, progress to facultative and then finally to obligate symbiotic relationships (Moya et al. 2008). At the terminus of this progression, we have obligate endosymbiotic relationships (mutualisms) where neither individual can survive independently, for instance, the pea aphid Acyrthosiphon pisum and its endosymbiont Buchnera aphidicola (Scarborough et al. 2005; Shigenobu and Wilson 2011), whereas legumes and rhizobial bacteria, a facultative commensalism, sit earlier in the continuum. Current models of symbiosis are heavily skewed towards the terminus of this evolutionary progression, obligate mutualistic endosymbiosis. However, many symbioses in the marine environment fall early in the continuum, and, while some of these relationships progress to obligate interactions, many are maintained at the casual or facultative level (Fisher et al. 2017). Thus, important questions arise: how do symbioses evolve from casual to facultative to dependent relationships? When does symbiosis begin (Figure 1.1)?
Figure 1.1 Defining the symbiosis space. X-axis represents the level of interdependency between host and symbiont from casual to facultative to obligate. Y-axis represents the length of the interaction from brief to whole lifecycle. Dotted arrow represents a possible trajectory from free-living to endosymbiotic state.
1.2 Chemical cues mediate symbiotic interactions
The successful establishment of many classic models of horizontally acquired symbiosis (Euprymna-Vibrio; legumes-Rhizobia/mycorrhizal fungi) depend upon successful and specific signal exchanges between partners. Biologists have begun to decipher the chemical languages and cellular and molecular events involved in these exchanges revealing them to be multistage events of some complexity. The complexity of these exchanges strongly suggests that (in their current form) they almost certainly postdate the evolutionary start of the relationship, and likely have limited value in discovering how symbioses develop initially. Furthermore, in the marine environment, most host organisms first interact with multispecies bacterial biofilms as larvae (Lema et al. 2019). How are the “right” signals heard amongst the noise? We argue that examination of pre-symbiotic bacteria–eukaryotic interactions in non-model organisms will greatly enhance our understanding of the interactions and ecological drivers that establish and maintain symbioses.
1.3 How do specific symbiotic interactions begin? Examples from the pre-symbiosis space
Inter-kingdom associations that exist early in the symbiosis continuum (Figure 1.1) can inform our understanding of how symbioses develop. Bacteria can interact with eukaryotes in a number of ways that are not classically defined as symbiosis but result in modifications to the eukaryote’s physiology or behavior. While not strictly symbioses, these interactions can be viewed as existing on the edge of the symbiotic continuum space defined above. For example, rosette (colony) formation and sexual reproduction in choanoflagellates are induced and inhibited by different bacterial metabolites (Levin and King 2013). Rosette formation in Salpingoeca rosetta is induced by a combination of lipid molecules from the prey bacterium Algoriphagus machipongonensis (Alegado et al. 2012; Beemelmanns et al. 2014; Woznica and King 2018). In this example, one form of eukaryotic development is dependent on the presence of appropriate prey microbes; however, the microbes themselves experience only the negative selection pressure of being eaten. Although not a symbiosis, the choanoflagellate example demonstrates how interactions with bacteria can not only drive eukaryotic development but also reveal potential drivers for multicellularity.
Marine sponges and corals provide another valuable space in which to examine not only the interplay between host eukaryotes and their associated bacteria, but also how symbionts can be found amongst the “noise” of diverse bacterial populations. Marine sponges contain diverse consortia of bacteria: currently at least 39 microbial phyla have been demonstrated to associate with sponges (Pita et al. 2018). Additionally, the density of those bacteria can reach 109 microbial cells/cm3 of sponge (Hentschel et al. 2006). The coral microbiome is also a complex consortium: up to 69 phyla have been identified in association with stony corals (Pollock et al. 2018; Huggett and Apprill 2019). In contrast to sponges and corals, most other animals interact with only 3–5 bacterial phyla (Kostic et al. 2013). The natural variability of both microbes and symbioses is reflected in the variability of interactions, from transients to true symbionts, and positive to negative (Figure 1.1). While unravelling the complex mechanisms that allow hosts to identify friend from foe amongst such complex communities remains experimentally challenging (Pita et al. 2016), a number of commonalities are emerging, for example, core groups have been identified that associate with taxonomic consistency (Rohwer et al. 2002; Erwin et al. 2012; Hester et al. 2015; Ainsworth and Gates 2016; Bourne et al. 2016; Thomas et al. 2016; Ainsworth et al. 2017). Many of the identified core microbiomes of these groups have ...
Table of contents
Cover
Half Title
Series Page
Title Page
Copyright Page
Contents
Series Preface
Preface
Contributors
Chapter 1: When does symbiosis begin? Bacterial cues necessary for metamorphosis in the marine polychaete Hydroides elegans
Chapter 2: The language of symbiosis: Insights from protist biology
Chapter 3: Trichoplax and its bacteria: How many are there? Are they speaking?
Chapter 4: Decoding cellular dialogues between sponges, bacteria, and phages
Chapter 5: Symbiotic interactions in the holobiont Hydra
Chapter 6: Hydra and Curvibacter: An intimate crosstalk at the epithelial interface
Chapter 7: The coral holobiont highlights the dependence of cnidarian animal hosts on their associated microbes
Chapter 8: Extra-intestinal regulation of the gut microbiome: The case of C. elegans TGFβ/SMA signaling
Chapter 9: Multiple roles of bacterially produced natural products in the bryozoan Bugula neritina
Chapter 10: The molecular dialogue through ontogeny between a squid host and its luminous symbiont
Chapter 11: Evolving integrated multipartite symbioses between plant-sap feeding insects (Hemiptera) and their endosymbionts
Chapter 12: Symbiosis for insect cuticle formation
Chapter 13: Microbial determinants of folivory in insects
Chapter 14: Right on cue: Microbiota promote plasticity of zebrafish digestive tract
Chapter 15: Uncovering the history of intestinal host–microbiome interactions through vertebrate comparative genomics
Chapter 16: Molecular interactions of microbes and the plant phyllosphere: The phyllosphere-microbiome is shaped by the interplay of secreted microbial molecules and the plant immune system
Chapter 17: Cellular dialogues between hosts and microbial symbionts: Generalities emerging
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Cellular Dialogues in the Holobiont by Thomas C. G. Bosch, Michael G. Hadfield, Thomas C. G. Bosch,Michael G. Hadfield in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.