Over the past forty years much work has assessed how attention modulates perception, but relatively little work has evaluated the role of attention in action. This is despite the fact that recent research indicates that the relation between attention and action is a crucial factor in human performance. Attention in Action provides state-of-the-art discussion of the role of attention in action and of action in constraining attention. The research takes an interdisciplinary approach covering experimental studies of attention and action, neuropsychological studies of patients with impaired action and attention, single cell studies of cross-modal links in attention and action, and brain imaging studies on the underlying neural circuitry. Contributions from prominent international researchers both review the field and present new evidence, making this book an invaluable resource for researchers and therapists alike.

- 384 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Part I
Functional processes
1 Attending to what you are doing
Neuropsychological and experimental evidence for interactions between perception and action
Glyn W. Humphreys, M. Jane Riddoch, Karina J. Linnell, David J. Punt, Martin G. Edwards and Alan M. Wing
Abstract
What are the relations between perceptual selection (e.g. for object identification) and action selection (the selection of an appropriate action to an object)? We discuss four pieces of novel neuropsychological and experimental evidence indicating that perception and action interact in determining human performance. First, we show that action relations between stimuli constrain the amount of visual extinction present in neuropsychological patients. There is less extinction when objects are in the correct co-locations for action. In addition, action relations between objects can lead to the coupling of movements to objects in patients who otherwise show uncoupled actions to separate objects (in Balint’s syndrome and in a case of motor extinction). Thus action relations between stimuli affect both perceptual selection (in cases of extinction) and action selection (in Balint’s syndrome and motor extinction). Subsequently, we note evidence indicating that (1) action relations between objects and (2) actions to objects combine multiplicatively in their effects on visual selection. Finally, we introduce evidence from normal observers showing that action to an object can change the nature of the processes mediating visual selection. Perception and action are coupled when there are action relations between objects in the environment and when actions are made towards stimuli.
Perceptual selection and selection for action
Over the last 10 years or so, a good deal of evidence has accumulated that perception and action can dissociate from one another (for reviews, see Milner & Goodale, 1995; Rossetti & Pisella, 2002). For example, damage to different brain regions can selectively disrupt either perception but not action to visual stimuli (e.g. after damage to ventral areas mediating visual processing), or action but not the perception of stimuli (e.g. after damage to more dorsal areas of cortex). Similarly, the effects of some visual illusions can be larger on perceptual judgements than on actions (Aglioti, DeSouza, & Goodale, 1995; Bridgeman, 2002; Haffenden & Goodale, 1998; for alternative views, see Franz et al., 2000; Pavani et al., 1999), while effects of temporal delay can be larger on action than on perceptual judgement tasks (Milner & Goodale, 1995). This contrast between perception and action may extend also to the processes involved in selecting between multiple stimuli for perception and in selecting the appropriate actions to a stimulus (selection for perception and selection for action). Neuroanatomical research indicates that selection for perception and selection for action to visual stimuli depend, respectively, on networks within anterior (frontal) and more posterior (parieto-occipital) regions (e.g. Posner & Petersen, 1990). Neuropsychological research indicates that patients with parietal damage can be impaired in the perceptual selection of stimuli on one side of space, rather than in acting to the affected side. For example, when required to cancel lines seen through a mirror, patients can have problems in orienting visual attention towards stimuli on the perceived contralesional side of space, even when an action to that stimulus would be made on the ipsilesional side. In contrast, they may orient appropriately to stimuli on the perceived ipsilesional side even when their actions are then made on the contralesional side (e.g. Ackroyd et al., 2002; Na, Adair, Williamson, Schwartz, Haws, & Heilman, 1998). This suggests a problem in perceptual selection rather than in acting towards the affected side. In contrast, patients with damage to, or disconnection of, more frontal brain regions can be impaired in selecting a task-appropriate action to a stimulus even though they select the appropriate stimulus to make an action to (e.g. Riddoch, Humphreys, & Edwards, 2000a, 2000b). In this case, action selection seems to be impaired while perceptual selection continues to operate. On this view, there are separate systems not only for object identification and the parameretization of actions to objects (e.g. Milner & Goodale, 1995), but also for selecting the objects to identify and the actions to make in the first place.
Dissociations between tasks are useful for highlighting structural constraints on information processing, since they indicate that the processing system must have an anatomical and functional organization such that one task can be affected more than another. However, this does not rule out the possibility that processes that are isolatable under some circumstances nevertheless interact to determine normal performance. Indeed, there is mounting evidence that perception can be affected by action. For example, our ability to identify briefly presented stimuli is tightly bound to where an action is made. Deubel, Schneider and colleagues (e.g. Deubel & Schneider, 1996, this volume; Deubel, Schneider, & Paprotta, 1998; Schneider & Deubel, 2002) have presented elegant evidence that identification is improved if we point to a location where a stimulus appears, relative to when we point to another location close by. This appears to be a necessary coupling, because the same result occurs even when the stimulus is presented at a constant location; if perceptual selection could normally be insulated from effects of action, then we ought to be able to set perceptual selection to the known stimulus location independent of where an action is directed to. In addition, perceptual judgements (such as whether stimuli are inverted or not) are affected by the position of the handle of the stimulus with respect to the effector used for the response and by compatibility between the particular response required (fine vs power-grip) and the response typically used with the object (Ellis & Tucker, 2000; Tucker & Ellis, 1998, 2001). This suggests that perception is influenced by action-related properties of objects (e.g. the degree to which an object “affords” a particular action; cf. Gibson, 1979; see Phillips & Ward, 2002, for an alternative account of some of these last results). Even in frontal-lobe patients showing dissociations between (impaired) action selection and (spared) perceptual selection, there can be incorrect selection of a stimulus for action if a distractor has the same perceptual attribute as that of a stimulus that was recently responded to and if the distractor carries a strong affordance (Boutsen & Humphreys, 2003). Here a temporary association between the perceptual attribute recently used for selection, the affordance of the stimulus and the response last made seems to increase the potency of that attribute for subsequent perceptual selection. Thus perception appears to be sensitive to action-related properties of stimuli, while perceptual selection is influenced by where an action is directed to and by temporary associations between perceptual attributes, object affordances for action and the recent response history. Perceptual selection and action selection may interact in determining both the object to which an action is directed and the action that is used.
In this chapter, we review recent neuropsychological and experimental evidence from our laboratory supporting the view that perceptual selection and action selection are interactive processes. The evidence is also relevant to the form of the interactions between the processes. For example, we suggest that action relations between objects in the environment play an important role in both perceptual selection and action selection, enhancing any effect of action towards an object. In addition, we propose that making an action to an object does not merely add to the processes that determine perceptual selection (e.g. increasing the activation of targets that are visually attended) but it changes the way they operate. We begin by first discussing the effects of action relations on perceptual selection and on action.
Action relations affect perceptual selection: evidence from visual extinction
There is a large amount of experimental and neuropsychological evidence that perceptual selection is influenced by factors that affect object recognition, such as whether visual elements group on the basis of a Gestalt property (e.g. continuation, collinearity, common motion), whether they form a known object, or whether separate objects are associatively related. Particularly clear examples come from the neuropsychological syndrome of visual extinction, where patients (often with unilateral damage involving the parietal lobe) are good at selecting a single stimulus presented on the contra-lesional side of space but are unable to perceive the same item when another (independent) stimulus appears simultaneously on the ipsilesional side (e.g. Karnath, 1988). Extinction can decrease, however, if the contra-and ipsi-lesional stimuli group by continuation, collinearity, common enclosure, common shape and contrast polarity (e.g. Gilchrist, Humphreys, & Riddoch, 1996; Humphreys, 1998; Mattingley, Davis, & Driver, 1997; Ward, Goodrich, & Driver, 1994). Even when low-level Gestalt factors are unavailable to link stimuli, extinction can be reduced if stimuli are parts of a known object (e.g. if two letters form a word rather than a nonword; Kumada & Humphreys, 2001). It can also be reduced if two stimuli are associatively related to one another (e.g. with two associated words; see Coslett & Saffran, 1991). The factors that are influential here (Gestalt grouping, object unit formation and associative relations between stimuli) can all be conceptualized as operating within a system that jointly recognizes objects and selects between competing perceptual representations.
In addition, are the competitive processes underlying perceptual selection also sensitive to action relations between stimuli (e.g. whether or not two stimuli are in the appropriate locations to be used together)? Furthermore, are any effects of action relations dissociable from whether or not stimuli are associatively related or form a familiar unit for object recognition? Recently, we addressed these questions in studies of patients with visual extinction, where we assessed the effects of action relations between stimuli on perceptual selection (Riddoch, Humphreys, Edwards, Baker, & Willson, 2003). Five patients were tested, three with unilateral right parietal damage (JB, MB, MP), one with unilateral left parietal damage (RH) and one with bilateral parietal damage, more pronounced on the right than on the left (GK). All the patients showed spatial extinction, with items on the contra-lesional side of space being difficult to identify when another item appeared simultaneously on the ipsilesional side.1 In a first experiment (Experiment 1), we assessed the identification of pairs of objects that were commonly used together and that were either placed in the correct spatial locations for actions (e.g. a corkscrew going into the top of a wine bottle) or placed in incorrect spatial locations for action (the corkscrew going into the bottom of the bottle; see Figure 1.1 a). In both the correct and incorrect location conditions, the pairs were presented twice, with each object appearing once in each field. In addition, single-object trials were included in which eachindividual stimulus was exposed in the same location as it appeared in the two-object trials. The task was to identify the objects presented on a trial, and stimulus exposure times were adjusted for individual patients so that they were able to identify about 80% of the contralesional objects on single-item trials (Figure 1.2 a). On two-object trials, performance was reliably better when the stimuli were placed in the correct positions for action relative to when they fell in incorrect positions for action (Figure 1.2b). This held across all the individual patients. This effect was not due to guessing. First, the same objects were used in the correct and incorrect location conditions, so the chances of guessing the identity of the contralesional item from the ipsilesional one should have been equal. In addition, while some of the patients reported that they could tell that “something” was present along with the ipsilesional item on unsuccessful two-object trials, two patients (MP and GK) made errors in which they reported that only an ipsilesional item occurred, but they never made errors on single-item trials by guessing the identity of a second object. Thus is appears that perceptual selection inthe patients was better if two objects fell in the correct positions for action relative to when they were in incorrect positions.
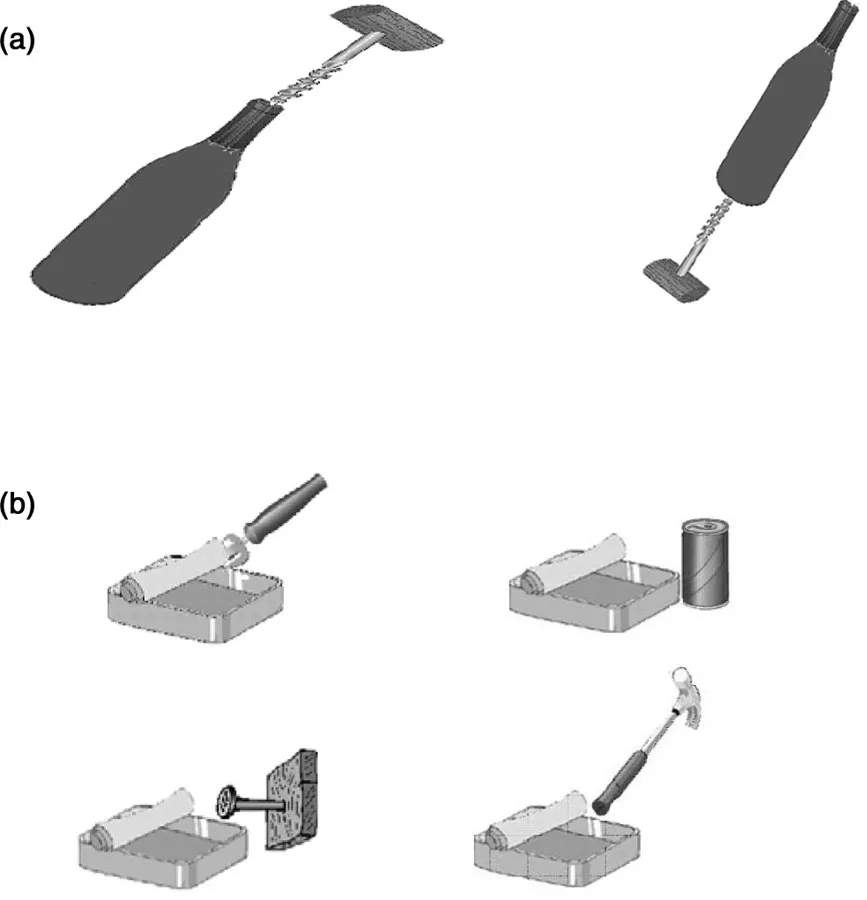
Figure 1.1 Example stimuli from studies examining the effects of action relations between stimuli on visual extinction (after Riddoch et al., 2003). (a) The stimuli used in Experiment 1 contrasting report when objects are in correct versus incorrect positions for action. (b) The stimuli employed in Experiment 2 contrasting report with objects that are used jointly in action versus objects that are verbally associated.
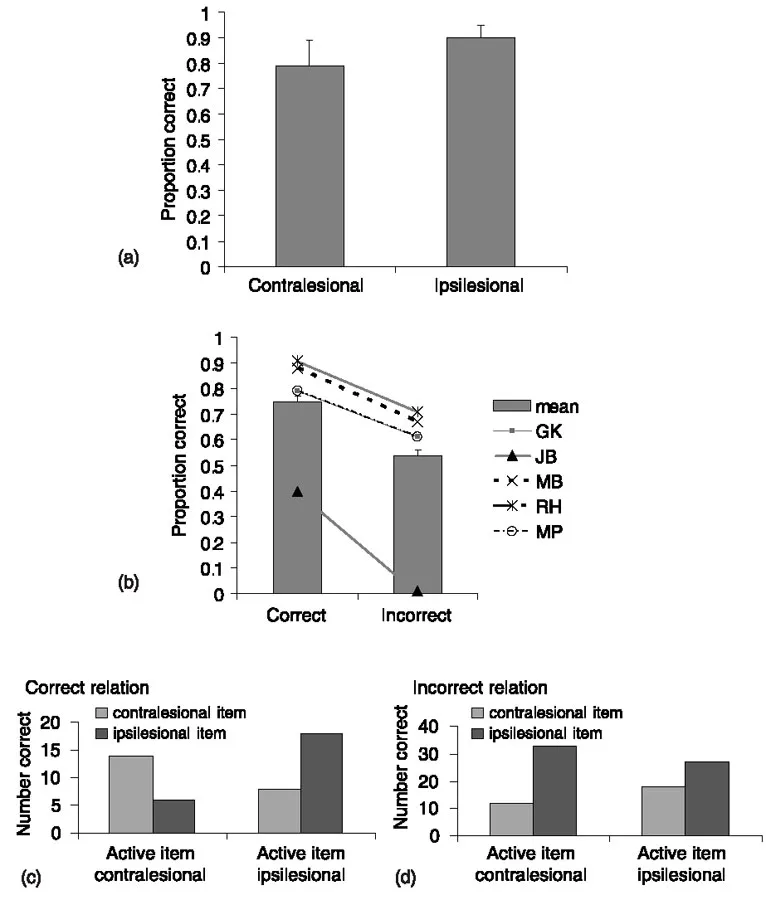
Figure 1.2 The results from Experiment 1. (a) Single-item reports. (b) Number of correct reports of two objects. (c, d) Data from trials on which only one object was reported (on two-item trials), as a function of whether the objects were in the correct (c) or incorrect (d) positions for action (after Riddoch et al., 2003).
We also examined the performance of the patients when they only reported one of the two objects present in a pair. We categorized each pair according to whether an object was an “active” partner (that was moved when the action was performed) or a “passive” partner (that was stationary during the action). In the example in Figure 1a, the corkscrew was categorized as the active partner and the wine bottle as the passive partner. A clear difference emerged between the identification of the active and passive partners, on trials where only one was reported, in the correct and incorrect location conditions. In the correct location condition, the patients tended to identify the active member of a pair irrespective of whether it was in the contra-or ipsilesional field. However, when the objects fell in incorrect locations for action, the patients tended to report the ipsilesional item irrespective of whether it was the active or passive partner in the pair (see Figure 1.2 c, d). This result is interesting, since it suggests that having...
Table of contents
- Cover Page
- Title Page
- Copyright Page
- Contributors
- Preface
- Part I functional processes
- Part II neural processes
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Attention in Action by Glyn Humphreys,Jane Riddoch in PDF and/or ePUB format, as well as other popular books in Psychology & Cognitive Neuroscience & Neuropsychology. We have over 1.5 million books available in our catalogue for you to explore.