
- 676 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Isolation and Purification of Proteins
About this book
This publication details the isolation of proteins from biological materials, techniques for solid-liquid separation, concentration, crystallization, chromatography, scale-up, process monitoring, product formulation, and regulatory and commercial considerations in protein production. The authors discuss the release of protein from a biological host, selectivity in affinity chromatography, precipitation of proteins (both non-specific and specific), extraction for rapid protein isolation, adsorption as an initial step for the capture of proteins, scale-up and commercial production of recombinant proteins, and process monitoring in downstream processing.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
Pharmacology1
Release of Protein from Biological Host
Rajni Hatti-Kaul and Bo Mattiasson
Lund University, Lund, Sweden
1. BIOLOGICAL HOSTS FOR PROTEIN PRODUCTION
The means to gain access to the product from a biological source is the primary consideration during downstream processing of proteins. Traditionally, the choice of a protein source was restricted to the natural biological material producing it in sufficient amounts. But recombinant DNA technology has completely changed the scenario by allowing protein production in a biological host of choice. Microorganisms clearly constitute the most common production systems for industrial enzymes and other proteins. Mammalian cell hosts are preferred only when correct posttranslational modification is essential for the function of eukaryotic proteins. Insect cells are also emerging as production systems; however, the posttranslational processing differs from the mammalian cell systems.
Despite the advantages of microorganisms as protein source, several protein products are still economically produced from animal and plant materials possibly because of sufficiently high amounts of these products in such sources. Animal tissues and organs provide excellent sources for some lipases, esterases, proteases, and other proteins. Hen egg white continues to be a good source for lysozyme. Cultivated plants provide adequate source for proteases like bromelain and ficin. Moreover, transgenic animals and plants are also appearing as commercial production hosts for heterologous proteins, the former being preferred for therapeutic proteins and the latter for industrial enzymes.
Access to the product is simple and inexpensive when the protein is produced extracellularly. Separation of biomass or any other particulate matter is the only requirement for obtaining the protein in a clarified form that is processed further for concentration and purification. Bulk enzymes are invariably produced extracellularly by Bacillus species and fungi, as are the proteins produced by mammalian cell culture. Protein production in transgenic animals is often directed to milk; here the clarification step involves the removal of lipids and casein.
Proteins associated with the cells, on the other hand, need to be released prior to commencement of downstream processing. The method of protein release is often determined by the kind of host cells/tissue harboring it and its exact location. Most cell-associated proteins are located in the cytoplasm, the soluble fraction of the cells, whereas some are present in the plasma membrane enveloping the cytoplasm and others in the periplasmic space separating the membrane from the cell wall. In Escherichia coli, which is often the microbial host of choice for production of recombinant proteins, the foreign protein normally accumulates in the cytoplasm either in the soluble form or, in the case of overproduction, as insoluble inclusion bodies. Saccharomyces cerevisiae is another industrial host particularly when a GRAS (generally regarded as safe) organism is required for a particular process, and its secretory mechanism does not function for all proteins that are usually produced as soluble intracellular products.
Different types of cells vary basically in the wall structure that provides elasticity and mechanical strength to the cells, and hence in their resistance to cell breakage. Cell wall is lacking in animal cells, which are thus easy to break. In gram-positive bacteria, peptidoglycan constitutes the major cell wall component associated with teichoic acids and polysaccharides, whereas in gram-negative bacteria, the peptidoglycan structure is covered by another wall layer composed of proteins, phospholipids, lipopoteins, and lipopoly-saccharides (Fig. 1). There are also substantial variations in the structure among different bacterial species, and also in the presence of capsules and slime layers, that further influence the resistance of cells to disruption. The cell wall in yeasts is made of glucans, mannans, and proteins, which form a highly cross-linked structure (Fig. 1), whereas in a typical mature plant cell it is composed of cellulose and other polysaccharides.
2. GENETIC APPROACHES FOR EXPORT OF RECOMBINANT PROTEINS
In order to avoid the cell disruption stage and the problems related to subsequent processing, various strategies have been attempted at the genetic level to make the cells secrete the protein. Expression of proteins in peri-plasm is also preferred since the amount of contaminating proteins is significantly lower than in the cytoplasm. Furthermore, because of its oxidizing environment the periplasm constitutes an attractive destination for the production of eukaryotic proteins (Baneyx 1999). Polypeptides destined for export are synthesized as preproteins containing a signal sequence 18–30 amino acids long at the amino terminus, which is cleaved during the translocation by inner-membrane-associated signal peptidases. Many signal sequences derived from naturally occurring secretory proteins (e.g., OmpA, OmpT, PelB, β-lactamase, and alkaline phosphatase) allow the translocation of heterologous proteins across the inner membrane when fused to their amino-terminal ends (Baneyx 1999; Pines and Inouye 1999).
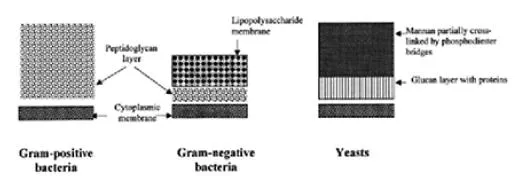
Figure 1 Schematic presentation of cell envelopes of bacteria and yeast.
Excretion of human growth hormone to the culture medium of E.coli cells was reported using excretion vector (pEAP8) (Kato et al. 1987). The passage through the bacterial inner membrane was made possible using the penicillinase signal sequence of an alkalophilic Bacillus sp. whereas the outer membrane was rendered permeable by the insertional activation of the kil gene (derived from plasmid pMB9) (Kobayashi et al. 1986). In another study, induction of β-lactamase synthesis from a strong tac promoter resulted in the release of more than 90% of the total soluble enzyme activity into the medium (Geourgiou et al. 1988). Excretion was not significant when the promoter was not fully induced or when the enzyme synthesis was directed by a weaker promoter. Permeability of the outer membrane of the excreting cells appeared to be affected as seen by higher sensitivity to detergents and also by release of periplasmic enzymes into the medium.
Abrahamsen et al. (1986) reported the extracellular secretion in E.coli of insulin-like growth factor I as a fusion protein with a protein A fragment composed of two IgG-binding fragments (EE). The presence in the protein A gene of a stress-induced promoter functional in E.coli was suggested.
3. RELEASE OF CELL-BOUND PROTEIN
The different strategies that may be used for releasing the cell-bound proteins include breaking the cells structure by mechanical forces (e.g., homogenisation or grinding with abrasives), damaging preferentially the cell wall (e.g., by drying or enzymatic lysis), or lysing primarily the membranes (e.g., by treatment with chemicals). Different mechanical and nonmechanical methods of disruption of microbial cells have been reviewed earlier in several papers (Harrison 1991; Schütte and Kula 1993; Chisti and Moo-Young 1986; Middelberg 1995). For most of the methods used, a number of factors, including type of organism, cultivation conditions, storage conditions, growth phase, viscosity and rheological properties of the cell suspension, pH, and so forth, influence the outcome of the disruption. Hence, expressing proteins into standard organisms like E.coli and yeasts that have established disruption procedures minimizes the extent of optimization required for cell breakage.
Development of a cell disruption process cannot be isolated from further downstream processing because the operation affects the physical properties of the suspension, such as viscosity, density, particle size, and settleability, which influence the subsequent solid-liquid separation step (Chisti and Moo-Young 1986). The product release can also be accompanied by its denaturation, due to the stress forces generated by the procedure used to rupture the cells, digestion by the proteases liberated from the cells, or interaction with other nonprotein constituents of the cells. A practical consideration of importance when evaluating the current methods available for industrial scale breakage of microbial cells is to ensure the availability both of small-scale equipment for development and optimization of the process, and of equipment of suitable capacity for the proposed largescale operations.
3.1 Mechanical Disruption of Microbial Cells
Mechanical disruption is the most common mode for protein release from cells despite the higher capital and operating costs, stress to the proteins, and generation of viscous homogenate with fine cell debris as compared with the nonmechanical methods. The disruption is based primarily on liquid or solid shear forces.
The liquid shear cell disruption is often associated with the cavitation phenomenon that involves formation of vapor cavities in liquid due to local reduction in pressure that could be affected by ultrasonic vibrations, local increase in velocity, and so forth. Collapse and rebound of the cavities will occur until an increase in pressure causes their destruction. On the collapse of the cavitation bubble, a large amount of energy is released as mechanical energy in the form of elastic waves that disintegrate into eddies. According to Doulah (1977), the eddies larger than the dimension of the cell will move it from place to place whereas the smaller eddies will impart motions of different intensities to the cell, creating a pressure difference across the cell. When the kinetic energy content of the cell exceeds the cell wall strength, the cell disintegrates. Cavitation also produces free radicals that lead to protein denaturation (Save et al. 1997).
An established technique for cell disruption in small samples that involves cavitation is ultrasonication, i.e., using ultrasonic vibrations covering a frequency range extending upward from 20 kHz (Chisti and Moo-Young 1986; Harrison 1991). This technique is not used at industrial scale primarily because the ultrasonication energy absorbed into suspensions ultimately appears as heat, and good temperature control is necessary. On the other hand, hydrodynamic cavitation results from pumping of a cell suspension through a downstream constriction, which causes a local increase in velocity (Save et al. 1994, 1997; Harrison 1991). Disruption of yeast (Save et al. 1994, 1997) and Alcaligenes eutrophus (Harrison 1991) by such a process was shown to be more energy efficient than the ultrasonic cavitation. However, in contrast to ultrasonication, the disruption was influenced significantly by the concentration of cells.
The established large-scale cell disruption operations based on liquid and solid shear are high-pressure homogenization and bead milling, respectively, with the former being the more common of the two. These will be described in some detail here. Much of the industrial disruption equipment has been inherited from chemical and food processing industries. Although no radical changes have occurred in cell disruption technologies over the years, major efforts have been put into introducing sterilization and containment as secondary features for biotechnology applications in order to ensure safe operation with recombinant organisms (Foster 1995).
High-Pressure Homogenization
A high-pressure homogenizer comprises a positive displacement pump that delivers fluid at a relatively constant flow to a homogenizing valve (Engler 1990; Pandolfe 1993; Pandolfe and Kinney 1998). The fluid exits through an adjustable gap between the valve and the valve seat and ultimately impinges on an impact ring as seen in Fig. 2. The flow restriction in the valve assembly drives up pressure (in the range of 50 and 120 MPa), which is then drastically reduced as the high-velocity jet of fluid emerges from the opening. This process applied to a microbial cell suspension results in the disruption of cells. Disruption follows first-order process at a given pressure in a high-pressure homogenizer. The extent of protein release is represented by (Hetherington et al. 1971; Schütte and Kula 1993):

where Rm and R are the maximal amount of protein available for release and the protein amount released at a certain time, respectively (kg protein/ kg cells), k is the first-order rate constant, N the number of passages, P the operating pressure, and a the pressure exponent.
The values of the constants are contingent on the nature of the organism and its culture conditions. However, the cell concentration over a relatively broad range is seen to have little influence on the disruption (Hetherington et al. 1971). Studies have shown that specific enzymes are released at different rates depending on their location in the cell; the peri-plasmic enzymes at a rate faster than the total protein, all cytoplasmic enzymes at the same rate as total protein, whereas membrane-bound enzyme slower (Follows et al. 1971). High-pressure homogenization is preferred for disruption of nonfilamentous organisms since the mycelial organisms result in clogging of the homogenizing valve (Zetelaki 1969; Keshavarz et al. 1990a).
Figure 2 Flow path of a cell suspension through a homogenizing valve in a high-pressure homogenizer.
Optimization of the cell disruption process is achieved by variation in the operating pressure and valve design (Engler 1990, Schütte and Kula 1993). Increasing the operating pressure reduces the number of passes required for disruption and hence the operating costs (Engler 1990; Pandolfe 1993). It also indirectly reduces the severe downstream clarification problems arising due to the fine cell debris resulting from further disintegration of already disrupted cells on repeated passage through the machine. However, the rise in pressure is also accompanied by a linear increase in temperature (2.5°C per 10 MPa) that necessitates rapid cooling of the homogenized suspension. While maintaining the pressure, alteration in homogenizing valve geometry can make significant changes in the protein yield (Pandolfe 1993; Pandolfe and Kinney 1998), e.g., disruption of baker’s yeast was more efficient with “knife edge” valve seat than the flat type unit (Hetherington et al. 1971).
Several investigators have sought to understand the mechanism of cell disruption by high-pressure homogenizer (Brookman 1974; Engler 1990; Keshavarz Moore et al. 1990b; Middelberg 1995; Kleinig and Middelberg 1998). According to Brookman (1974), magnitude of pressure drop is the major causative mechanism, while other researchers (Keshavarz Moore et al. 1990b; Engler 1990) suggested the involvement of two mechanisms—one relating to homogenization zone within the valve unit and the other with the impingement in the exit zone. The latter theory was confirmed by recent studies of Lander and coworkers (2000) with polysaccharides as a model shear-sensitive compound, where a similarity in the breakage pattern in terms of molecular size and polydispersity was observed with the results obtained from fluid shear flows in capillary tubes. Their results indicated that breakage occurs primarily by fluid shear in the valve unit, with a contribution by cavitation occurring in the impingement section where the jet of fluid strikes the impingement plate and the bubbles collapse as a result of repressurization of this impact. The occurrence of cavitation was indicated by the detection of free radicals in a pressure range from 11 to 35 MPa (Shirgaonkar et al. 1998; Lander et al. 2000).
The traditional form of high-pressure homogenizer is the Manton-Gaulin APV type, for which special designs for biotechnology applications have been developed (Pandolfe 1993). A few other types of equipment are now available. Microfluidizer (Microfluidics, Newton, MA) is one in which the cell suspension is driven at a flow rate of 250–600 ml/min by an air-powered intensifier pump at constant pressure (20–159 MPa) into an interaction chamber where the stream is split into two, which pass through precisely defined fixed-geometry microchannels (100μm), and are later directed h...
Table of contents
- Cover Page
- Title Page
- Copyright Page
- Preface
- Contributors
- Introduction
- 1 Release of Protein from Biological Host
- 2 Chromatography: The High-Resolution Technique for Protein Separation
- 3 Selectivity in Affinity Chromatography
- 4 Genetic Approaches to Facilitate Protein Purification
- 5 Solid-Liquid Separation
- 6 Membranes for Protein Isolation and Purification
- 7 Precipitation of Proteins: Nonspecific and Specific
- 8 Protein Crystallization for Large-Scale Bioseparation
- 9 Extraction for Rapid Protein Isolation
- 10 Adsorption as an Initial Step for the Capture of Proteins
- 11 Fast Chromatography of Proteins
- 12 Novel Approaches to the Chromatography of Proteins
- 13 Scale-up and Commercial Production of Recombinant Proteins
- 14 Process Monitoring During Protein Purification
- 15 Protein Formulation
- 16 Regulatory Requirements, Cleaning, and Sanitization Issues in Protein Purification
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Isolation and Purification of Proteins by Rajni Hatti-Kaul,Bo Mattiasson in PDF and/or ePUB format, as well as other popular books in Medicine & Pharmacology. We have over 1.5 million books available in our catalogue for you to explore.