
eBook - ePub
The Fish Immune System: Organism, Pathogen, and Environment
- 380 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Fish Immune System: Organism, Pathogen, and Environment
About this book
This book comprehensively reviews the immunology of fish--their health, interactions between them and their pathogens, and the impact of both endogenous and environmental changes on these interactions. Leading authorities provide an essential foundation for the understanding of fish immunology and fish health.As fish are increasingly used as model systems for vertebrate immune systems, The Fish Immune System will be a crucial reference.
- The only comprehensive, single-volume reference on the fish immune system
- Contributions from an international team of experts
- Useful to researchers interested in fish health as well as professionals managing fish hatcheries, aquariums, and other facilities that must maintain healthy fish
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
Sciences biologiquesSubtopic
Physiologie1
Cells and Tissues of the Immune System of Fish
Agustín G. Zapata, Akira Chibá and Alberto Varas
I. Introduction: The Phylogenetic Position of Fish
The first fossils recognizable as vertebrates are the Ostracoderms. They corresponded to a type of Agnathan fish that, apart from the lack of jaws, showed more primitive features than the modern fish, including the lack of paired fins and the presence of only a single, dorsally-located nostril. Nevertheless, modern Cyclostomes (i.e., myxinoids and lampreys) share many of their anatomical characteristics.
Jawed fishes arose nearly 400 million years ago during the Silurian period. Their descendants, the bony fish (Osteichthyes) and cartilaginous fish (Chondrichthyes) are the predominant forms in the current seas and freshwaters (Fig. 1). These two groups of fishes diverged from distinct Placoderm lines during the late Silurian or early Devonian period. The cartilaginous fishes, which are morphologically very similar to their Devonian ancestors, represent in a sense the end of an evolutionary line. In contrast, the bony fish constitute a progressive group that includes two large subdivisions, the ray-finned fishes (Actinopterygii) and the fleshy-finned fishes (Sarcopterygii). A small group of the latter, the Crossopterygians, were directly ancestral to amphibians. Another group of Sarcopterygii, the Dipnoi or lungfish, possess various morphological and embryological similarities to amphibians but are not directly in the line of emergence of higher vertebrates.
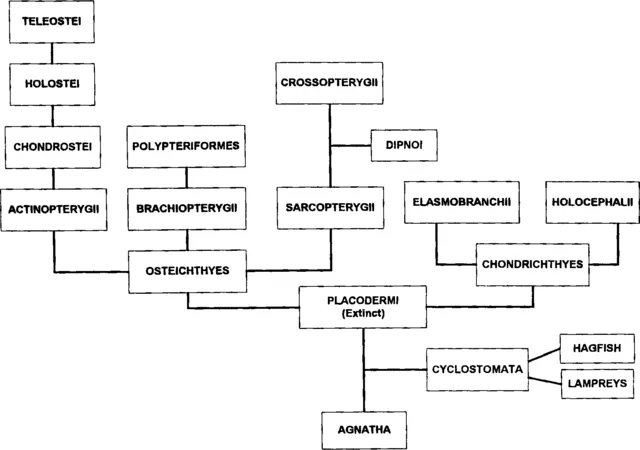
Fig. 1 A phylogenetical tree of fish.
II. Fish Leukocytes
Because several recent reviews have reported on the structural aspects of fish leukocytes in detail, and other chapters of this volume discuss fish blood cells, mainly monocytes and macrophages, we will describe only some of their more relevant features as an introduction to the principal aim of the chapter, the histophysiology of fish lymphoid organs.
A. Cyclostomes
In hagfish, only a single population of granulocytes has been reported (Tomonaga et al., 1973b; Mattisson and Fänge, 1977). These cells are probably heterophilic granulocytes of gnathostomes and constitute about half of the leukocytes in the blood of Myxine glutinosa (Mattisson and Fänge, 1977). They are spherical in shape with motile pseudopodia and contain a lobate nucleus. Their cytoplasmic granules are ovoid or rod shaped (Mattisson and Fänge, 1977). No peroxidase activity is found in the hagfish granulocytes (Johansson, 1973). These cells show amoeboid movement, are found extravascularly in various tissues, and at least in M. glutinosa, have phagocytic capacity (Mattisson and Fänge, 1977).
In contrast to the hagfish, different subpopulations of granulocytes are distinguishable in lampreys (Rowley et al., 1988). Among these subpopulations, the heterophilic granulocytes are found consistently and constitute about half of the leukocytes in the blood of adult Lampetra fluviatilis with a single subpopulation of the granulocytes. However, there are relatively low ratios of this subpopulation (8%) in the blood of the larval lampreys in which heterophilic granulocytes and acidophils are observed. On the other hand, acidophilic granulocytes are found in the blood of both larval and adult Lampetra spp and also both in the intestinal and renal lymphohemopoietic tissue of larval lampreys (Rowley et al., 1988). They are more common in the blood of ammocoetes than in adults. Basophilic granulocytes are reported only from Lampetra planeri (Fey, 1966) and Petromyzon marinus (Fey, 1966). Heterophilic granulocytes have a bilobed or trilobed nucleus and an azurophilic cytoplasm. Cytochemical studies show that they are acid phosphatase, esterase, and periodic acid-Schiff (PAS) positive, but peroxidase negative (Rowley et al., 1988). Electronmicroscopically, cytoplasmic granules are suggested to be in fact part of a single maturation series. In the lamprey, L. fluviatilis, heterophilic granulocytes phagocytose antibody-coated sheep erythrocytes in vitro (Fujii, 1981). The acidophilic granulocytes of P. marinus are round cells with minor cytoplasmic processes and contain an eccentric and irregularly shaped nucleus. Their cytoplasmic granules are electron dense, membrane bound, and homogeneous. Histo-chemical data on the peroxidase in the acidophilic granulocytes are few and inconsistent, for example, producing a negative reaction in Lampetra (Rowley et al., 1988) or variable reactions in L. planeri (Fey, 1966). Cytological characterization of the basophilic granulocytes is insufficient due to the paucity of data on this type of granulocyte.
B. Chondrichthyes
Little is known about holocephalan granulocytes (Hine and Wain, 1988a; Mattisson et al., 1990). Blood of Chimaera monstrosa contains two types of granulocytes referred to as coarse acidophilic granulocytes and fine granulocytes. The former has numerous coarse granules that stain bright red with Giemsa, whereas the latter is full of very fine, red granules. The granulocytes in the cranial lymphohemopoietic tissue also have been studied electronmicroscopically and have been designated as acidophils and heterophils. Cytoplasmic granules of acidophils are electron dense and homogeneous, whereas those of heterophils are heterogeneous in their internal structure (Mattisson et al., 1990).
Granulocytes of elasmobranchs have been studied in several species but their identification and terminology are somewhat confusing due to the enormous heterogeneity (Rowley et al., 1988). Mainwaring and Rowley (1985) reported four types of these cells in the blood of Scyliorhinus canicula and referred to them as G1, G2, G3, and G4 granulocytes. Of these cell types, G1, G3, and G4 are classified as acidophils and account for about 27, 3, and 9%, respectively, of the leukocyte population. G2 granulocytes, constituting 1.3% of the total leukocytes, correspond to heterophils. No basophilic granulocytes are found in the Scyliorhinus peripheral blood. In contrast, only two types of granulocytes, termed G1 and G2, were described in two species of rays, Raja clavata and R. microcellata. Both are categorized as acidophils. Among the Scyliorhinus granulocyte subpopulations, only G1 shows phagocytic activity and localization at sites of inflammation. Amoeboid activity and chemokinesis/chemotaxis to leukotriene-B4 are found not only for G1 but also G3 granulocytes. Strong acid phosphatase activity has been reported in G1, G2, and G3 granulocytes, but no peroxidase activity is demonstrated in any of the granulocyte subpopulations (Mainwaring and Rowley, 1985; Rowley et al., 1988). For more detailed information on cytological characteristics such as fine structure and developmental series of the elasmobranch granulocytes (see Ainsworth, ...
Table of contents
- Cover image
- Title page
- Table of Contents
- Inside Front Cover
- Front Matter
- Copyright page
- Contributors
- Preface
- Chapter 1: Cells and Tissues of the Immune System of Fish
- Chapter 2: The Nonspecific Immune System: Cellular Defenses
- Chapter 3: The Nonspecific Immune System: Humoral Defense
- Chapter 4: The Specific Immune System: Cellular Defenses
- Chapter 5: The Specific Immune System: Humoral Defense
- Chapter 6: Natural Changes in the Immune System of Fish
- Chapter 7: Environmental Factors in Fish Health: Immunological Aspects
- Chapter 8: Immunomodulation: Endogenous Factors
- Chapter 9: Infection and Disease
- Index
- Other Volumes in the Fish Physiology Series
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Fish Immune System: Organism, Pathogen, and Environment by William S. Hoar,David J. Randall in PDF and/or ePUB format, as well as other popular books in Sciences biologiques & Physiologie. We have over 1.5 million books available in our catalogue for you to explore.