
- 84 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Overflow Metabolism: From Yeast to Marathon Runners provides an overview of overflow metabolism, reviewing the major phenomenological aspects as observed in different organisms, followed by a critical analysis of proposed theories to explain overflow metabolism. In our ideal view of metabolism, we think of catabolism and anabolism. In catabolism nutrients break down to carbon dioxide and water to generate biochemical energy. In anabolism nutrients break down to generate building blocks for cell biosynthesis. Yet, when cells are pushed to high metabolic rates they exhibit incomplete catabolism of nutrients, with a lower energy yield and excretion of metabolic byproducts.
This phenomenon, characterized by the excretion of metabolic byproducts that could otherwise be used for catabolism or anabolism, is generally known as overflow metabolism. Overflow metabolism is a ubiquitous phenotype that has been conserved during evolution. Examples are the acetate switch in the bacterium E. coli, Crabtree effect in unicellular eukaryote yeasts, the lactate switch in sports medicine, and the Warburg effect in cancer. Several theories have been proposed to explain this seemingly wasteful phenotype. Yet, there is no consensus about what determines overflow metabolism and whether it offers any selective advantage.
- Includes examples of overflow metabolism and major phenomenological features
- Features a critical view of proposed theories to explain overflow metabolism
- Provides a summary of our preview work, proposing molecular crowding as the cause of overflow metabolism, together with new unpublished results
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
A Historical View of Overflow Metabolism
Abstract
Metabolism is the biological process where cells, tissues or whole organisms utilize organic molecules for survival, proliferation and to transform the environment. Metabolism is conceptually divided into catabolism and anabolism. Catabolism involves the breakdown of organic molecules to generate energy and precursor molecules for anabolism. Anabolism involves the synthesis of cell components from precursor molecules and energy. When catabolism is fully efficient, organic molecules are fully incorporated into the cell biomass or they are broken down to carbon dioxide to obtain the maximum energy yield. Under certain conditions, the organic molecules are only partially broken, and the products of incomplete catabolism are released to the environment. This phenomenon of incomplete catabolism, characterized by the excretion of metabolic products that could be otherwise incorporated in the cell biomass or catabolized to carbon dioxide, is called overflow metabolism.
Keywords
Catabolism; anabolism; overflow metabolism; Pasteur; Brown; Warburg; Crabtree
En résumé, la fermentation est un phénomène très-général. C'est la vie sans air, c'est la vie sans gaz oxygène libre
Louis Pasteur
1.1 Catabolism, Anabolism and Overflow
Metabolism is the biological process where cells, tissues or whole organisms utilize organic molecules for survival, proliferation and to transform the environment. Metabolism is conceptually divided into catabolism and anabolism. Catabolism involves the breakdown of organic molecules to generate energy and precursor molecules for anabolism. Anabolism involves the synthesis of cell components from precursor molecules and energy. When catabolism is fully efficient, organic molecules are fully incorporated into the cell biomass or they are broken down to carbon dioxide to obtain the maximum energy yield. Under certain conditions, the organic molecules are only partially broken, and the products of incomplete catabolism are released to the environment. This phenomenon of incomplete catabolism, characterized by the excretion of metabolic products that could be otherwise incorporated in the cell biomass or catabolized to carbon dioxide, is called overflow metabolism.
Glycolysis, the metabolic pathway comprising the first steps in the catabolism of sugars, is a classical example of the interplay between catabolism, anabolism and overflow metabolism (Fig. 1.1). Sugars are first broken down to glucose and glucose is then catabolized to two molecules of pyruvate. In its anabolic role, pyruvate is used to build larger molecules. Alternatively, pyruvate can be catabolized to carbon dioxide by the metabolic pathway of oxidative phosphorylation. The energy released by the catabolism of sugars is used to produce adenine triphosphate (ATP), the energy currency of cells. Glycolysis has a yield of 2 ATP molecules per molecule of glucose. Glycolysis combined with the oxidative phosphorylation of pyruvate has a much higher yield, which in mammalian cells amounts to 32 ATP molecules per molecule of glucose [1].
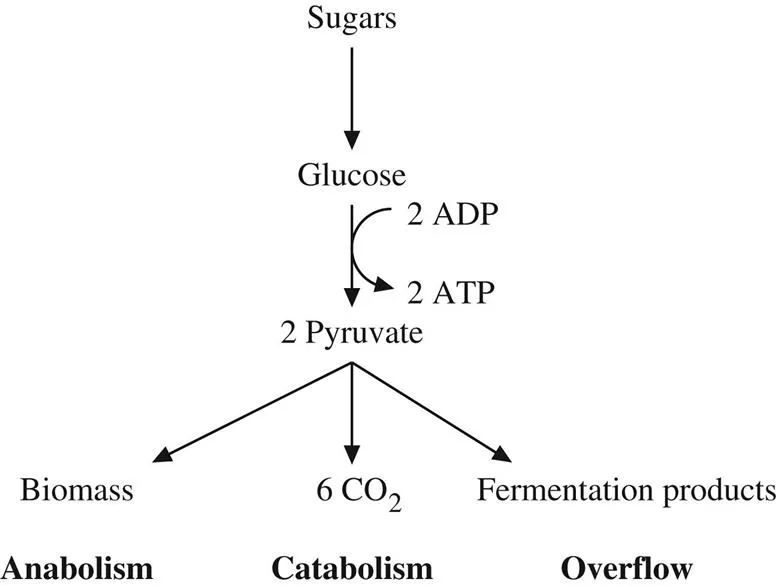
The fermentation of sugars to ethanol by baker’s yeast is a familiar example of overflow metabolism. Fermentation starts with the partial catabolism of sugars to pyruvate via glycolysis. However, instead of being further catabolised by oxidative phosphorylation, pyruvate is converted into an acid, gas or alcohol that is released from cells (overflow). The ATP yield of fermentation is that of glycolysis, 2 ATP molecules per molecule of glucose, 1/16 smaller than the ATP yield of oxidative phosphorylation.
1.2 Pasteur, Brown, Warburg and Crabtree
In 1861, Louis Pasteur found that aeration of yeast cultures caused yeast cell growth to increase and fermentation to decrease [2]. From those observations, Pasteur postulated that fermentation takes place in anaerobic conditions – ‘la vie sans gaz oxygene libre’ – and that oxygen has an inhibitory effect on fermentation [3] (Pasteur effect). The Pasteur effect was later recapitulated with muscle cells [4]. When resting muscle cells were maintained in a hydrogen atmosphere (and thus anaerobic) they produced lactate at high rates. In contrast, lactate production was almost entirely abrogated in an oxygen atmosphere.
The pioneering work of Pasteur pointed to the lack of oxygen as the cause of overflow metabolism in anaerobic fermentation. Some scientist contemporaneous with Pasteur questioned his theory, that fermentation was happening at the highest rate in the absence of oxygen. Adrian J. Brown, the half-brother of the more famous Horace T. Brown, conducted experiments to measure the rate of fermentation in the absence and presence of air [5]. In those experiments, yeast was added to two flasks, one with air flow blocked and another one exposed to air flow. When he analysed the amount of alcohol on each flask 3 h pass the start of fermentation, he observed the flask exposed to air contained more alcohol. Brown’s data indicated that fermentation is not restricted to anaerobic conditions. It can take place in the presence of oxygen at equal or higher rates. Brown was also the first to measure the rate of fermentation per cell in aerobic conditions [5]. Unfortunately, his work did not transcend his time, perhaps because of the wider acceptance of Pasteur hypothesis. I fortunately rediscovered his manuscripts when performing a Google search on the topic of aerobic fermentation. See Refs. [5,6] and a discussion of his work in Ref. [7].
It took 30 years for other biochemists to challenge the hypothesis that fermentation occurs at the highest rate in the absence of oxygen. In 1923, Otto Warburg and Seigo Minami reported that tumour tissues cultured in aerobic conditions manifested high rates of glucose fermentation to lactate, which were low in normal tissues [8]. A few years later, Crabtree reported the manifestation of aerobic fermentation by normal tissues infected with viruses [9].
The use of different experimental systems had a profound influence on the hypotheses Warburg and Crabtree pushed forward to explain aerobic fermentation. The observation of high rates of aerobic fermentation in cancer cells prompted Warburg to hypothesize that cancer cells have defective mitochondria [10,11]. That is a plausible extrapolation of the lack of oxygen as the cause of anaerobic fermentation. If fermentation occurs in the presence of oxygen, then there should be a defect in the cell machinery responsible for oxidative phosphorylation, the mitochondria in mammalian cells. In doing so, Warburg did not take into account the report by Crabtree of aerobic fermentation by normal tissues infected with viruses, which have competent mitochondria.
The recapitulation of aerobic fermentation in normal tissues subject to viral infection lead Crabtree to conclude that aerobic fermentation is not unique to cancer tissues [9]. Instead, and probably inspired by the formulation of the Pasteur effect as an inhibitory effect of oxygen on fermentation, Crabtree postulated that glycolysis has an inhibitory effect on oxidative phosphorylation: ‘Evidence is b...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Overflow Metabolism Wordle
- Dedication
- Preface
- Acknowledgement
- Chapter 1. A Historical View of Overflow Metabolism
- Chapter 2. Biochemical Horsepower
- Chapter 3. How Fast Can We Run?
- Chapter 4. How Fast Can We Grow?
- Chapter 5. Overflow Metabolism in Human Disease
- Chapter 6. Outlook
- References
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Overflow Metabolism by Alexei Vazquez in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biochemistry. We have over 1.5 million books available in our catalogue for you to explore.