- 322 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Neuroanatomy for Medical Students
About this book
Neuroanatomy for Medical Students, Second Edition provides a fundamental knowledge base that is essential to a proper understanding of the clinical neurosciences. This edition includes additional topics on neurophysiology, neuropharmacology, and applied anatomy. The areas on cell membrane structure and function, motor control, muscle spindles, spinocerebellar tracts, reticular formation, striatal transmitters, and retinal neurons are updated. This book also expands the topics on pineal gland, pituitary tumors, split brain effect, visual cortex, neural plasticity, and barrel fields. The topography of ventricles and summary table of cranial nerve are likewise revised. Other materials covered include nerve growth factor, neural transplantation, dorsal column transection, cerebellar memory, and perivascular spaces. The neurotransmitters and neuromodulators, nuclear magnetic resonance, and position emission tomography are also discussed. This publication is a good reference for medical students intending to acquire knowledge of basic neurobiology.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Development and topography of the nervous system
The purpose of this chapter is to present a preliminary overall view of the central nervous system. Those who have not yet studied embryology may prefer to start with general topography on page 12. Initially there may seem to be much new terminology, but this unfamiliarity resolves as studies progress: the glossary at the end of the book provides an explanation of all the neuroanatomical and clinical terms used. Many of the features briefly mentioned here can only be fully understood after further description and dissection. Commonly used descriptive terms are rostral (towards the beak, or nose); caudal (towards the tail); ventral (towards the belly) and dorsal (towards the back). These terms are equally applicable throughout the animal kingdom.
Development
An understanding of development helps to explain the nervous system’s organization (Fig. 1.1). In the early embryonic disc, ectoderm overlying the newly-formed notochord thickens to form a midline neural plate. The edges of the neural plate become elevated as folds, creating a neural groove. Fusion of these folds extends caudally from the cervical region, creating a neural tube, with small openings, the neuropores, at its rostral and caudal ends, which close by the end of the fourth intrauterine week. Vertebral bodies develop around the notochord, which persists as the nucleus pulposus of intervertebral discs. (Incomplete closure of the caudal neuropore and defective development of associated vertebral arches produce spina bifida). The lateral margins of the neural plate comprise specialized neural crest cells; neural tube formation segregates these, and in the process of embryonic segmentation they become dorsal root ganglia. (Other neural crest cells provide neurolemmal sheath cells for spinal nerve fibres or migrate to become sympathetic ganglion cells and chromaffin cells of suprarenal medulla.) The rostral part of the neural tube enlarges into forebrain, midbrain and hindbrain vesicles; the remainder remains cylindrical as the spinal cord; neural proliferation in its walls eventually narrows the lumen to a minute central canal.
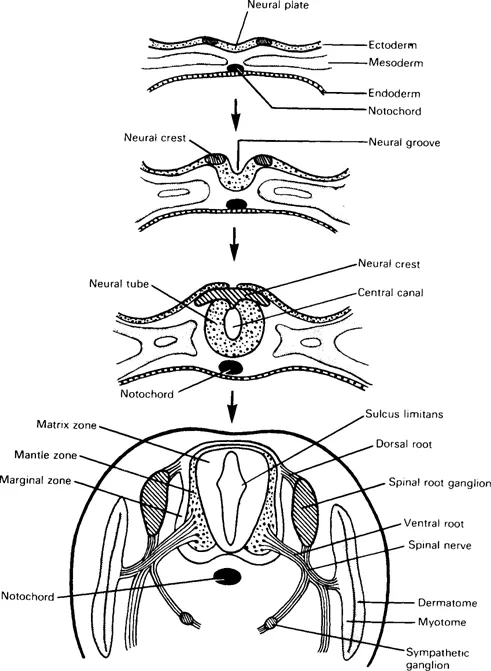
Fig. 1.1 Transverse sections showing progressive differentiation of the neural tube and associated structures.
The spinal cord
Details of histogenesis are beyond this brief account. Transverse sections of the neural tube reveal three layers (Fig. 1.1). The inner matrix zone is a wide germinal layer, its numerous cells undergoing mitosis; it produces neuroblasts and spongioblasts, the former developing into neurons, the latter into neuroglial cells (astrocytes and oligodendrocytes). The neuroblast cell bodies migrate outwards and form a surrounding mantle zone, the future spinal grey matter; their axons pass out further into a marginal zone, the future white matter. Central processes from the dorsal root ganglia grow into the neural tube; some ascend in the marginal zone, while others synapse with neurons in the mantle zone. When cell differentiation is complete, the residual cells of the matrix zone form the ependymal lining of the central canal.
The dorsal and ventral walls of the neural tube remain thin as roof and floor plates. On each side the wide mantle zone is demarcated into dorsal (alar) and ventral (basal) regions by an inner longitudinal sulcus limitans. Neurons of the alar lamina are functionally sensory (afferent). Neurons of the basal lamina are motor (efferent), their axons leaving the spinal cord as ventral roots which, with peripheral processes of the dorsal root ganglia, form spinal nerves (Fig. 1.2).
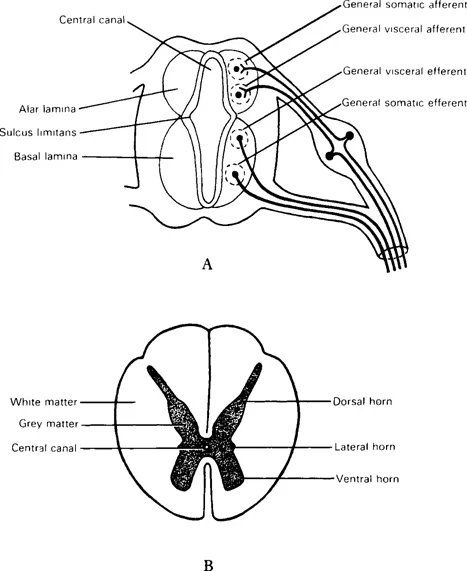
Fig. 1.2 Transverse sections of developing spinal cord showing (A) four cell columns in grey matter; (B) mature (thoracic) spinal cord.
Alar and basal laminae are subdivided into four longitudinal cell columns with specific functions. These grey columns are seen as ‘horns’ in cross-sections of a mature cord. The two afferent columns of each alar lamina receive axons from the dorsal root ganglia. Axons from the efferent columns form ventral nerve roots.
The general somatic afferent column (‘ordinary’ sensation) extends throughout the spinal cord and occupies most of the dorsal horn. It receives impulses from superficial (cutaneous) and deep (proprioceptive) receptors.
The general visceral afferent column (visceral sensation) at the base of the dorsal horn of cord segments T1–L2 and S2–4, receives impulses from viscera and blood vessels.
The general visceral efferent column (to smooth muscle) provides autonomic innervation for viscera, glands and blood vessels. Sympathetic outflow is from a lateral horn (T1–L2). Parasympathetic outflow is via certain cranial nerves and from S2–4 spinal segments. These fibres are termed ‘preganglionic’ because they all synapse in ganglia before reaching their targets.
The general somatic efferent column (to skeletal muscle) extends throughout the spinal cord in the ventral horn.
These four columns are termed ‘general’ because additional ‘special’ components are required in the brainstem for faculties such as taste and hearing. Aggregations of nerve cell bodies (somata), visible in transverse sections of grey matter, are often referred to as ‘nuclei’; each nucleus has particular functions and its neurons share common pathways.
Developmentally one cord segment (neuromere) serves one myotome and one dermatome on each side. In contrast to the very obvious segmentation of mesodermal somites, embryonic cord segments are not distinctly separated from one another because the developing cord must have internal structural continuity. Functional segmentation is marked externally by the attachments of pairs of spinal nerves.
Initially the spinal cord and vertebral canal are of equal length, but the former grows less rapidly; at birth its caudal end is level with the third lumbar vertebra and in adults reaches only to the disc between the first and second lumbar vertebrae. The more caudal spinal nerve roots are therefore elongated and pass obliquely within the canal before emerging via intervertebral foramina; beyond the spinal cord’s tip the vertebral canal contains a bundle of lumbar, sacral and coccygeal roots descending to their respective foramina.
Three membranes, derived from mesenchyme, surround the brain and spinal cord; these meninges are termed, from within outwards, pia mater, arachnoid mater and dura mater, and will be described later.
The brain
Three brain vesicles in the rostral part of the neural tube indicate the early division of the latter into forebrain (prosencephalon), midbrain (mesencephalon) and hindbrain (rhombencephalon) (Fig. 1.3); their cavities become ventricles in the mature brain. Three flexures appear in this region (Fig. 1.4); two are convex dorsally, a cephalic flexure (at midbrain level) and a cervical flexure (at the junction of hindbrain and spinal cord). A pontine flexure, concave dorsally, produced by unequal growth at future pontine level, has a buckling effect, everting the lateral walls and attenuating the roof of the neural tube here (Fig. 1.5 and see Fig. 5.4). The sensory alar laminae thus become lateral to the motor basal laminae in the floor of a rhomboid-shaped fossa (hence the name rhombencephalon). The part of the hindbrain caudal to the pontine flexure is the myelencephalon (future medulla oblongata); the rostral part, from which the pons and cerebellum develop, is the metencephalon; the hindbrain cavity becomes the fourth ventricle. In contrast, the mesencephalic cavity remains narrow as the cerebral aqueduct. The forebrain vesicle develops bilateral outgrowths which together constitute the telencephalon (= ‘end-brain’); these overgrow and cover the original forebrain, which becomes the diencephalon (= ‘between-brain’). The twin cavities of telencephalon develop into two lateral ventricles; the midline cavity of diencephalon is the third ventricle.
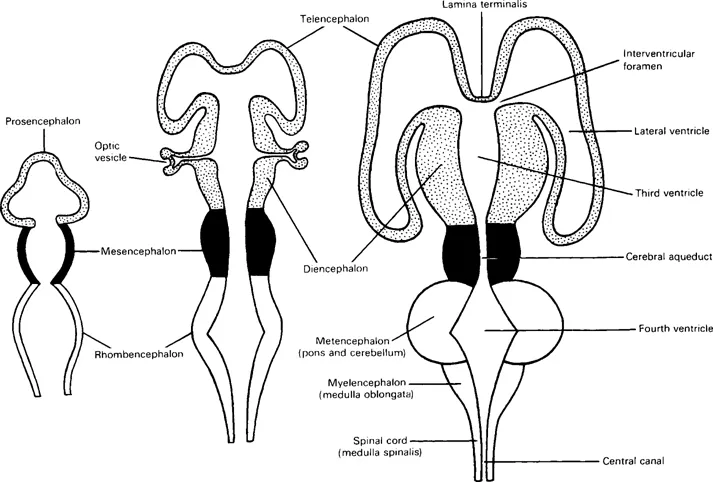
Fig. 1.3 Diagrams of stages in the differentiation of cerebral vesicles and the ventricular system.
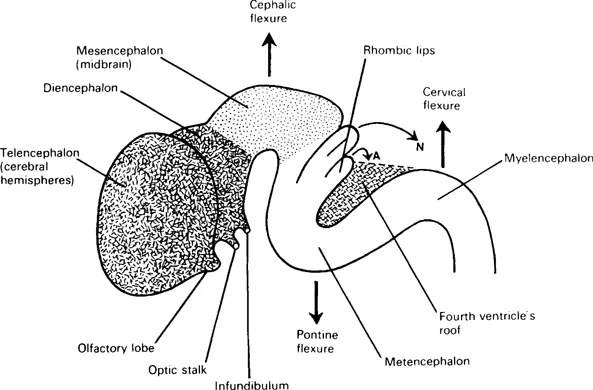
Fig. 1.4 Diagram of the external form of a devel...
Table of contents
- Cover image
- Title page
- Copyright
- Foreword
- PREFACE
- Chapter 1 Development and topography of the nervous system
- Chapter 2 Neurons and neuroglia
- Chapter 3 Peripheral nervous system
- Chapter 4 Spinal cord
- Chapter 5 Brainstem
- Chapter 6 Cranial nerves
- Chapter 7 Cerebellum
- Chapter 8 Diencephalon and internal capsule
- Chapter 9 Corpus striatum
- Chapter 10 Olfactory and limbic systems
- Chapter 11 Visual system
- Chapter 12 Cerebral cortex
- Chapter 13 Meninges, cerebrospinal fluid and cerebral ventricles
- Chapter 14 Blood supply of the central nervous system
- Chapter 15 Neurotransmitter pathways of the central nervous system
- Glossary: neuroanatomical and clinical terminology
- Bibliography
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Neuroanatomy for Medical Students by J. L. Wilkinson in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Human Anatomy & Physiology. We have over 1.5 million books available in our catalogue for you to explore.