Definitions
Endocrinology is the study of hormones; but what are hormones? The question is far more difficult to answer today than it was a few decades ago. The classic definition is that hormones are chemical substances produced by specialized tissues and secreted into blood, where they are carried to target organs. However, this definition was constructed when most of the available knowledge of endocrinology was restricted to vertebrate systems. As the field of endocrinology has expanded, new hormones and new systems that previously would not have been included under this definition have been discovered. It is useful to describe these discrepancies so that a more functional definition can be developed.
1. Specialized tissues for hormone synthesis. Discrete endocrine glands exist only in arthropods, mollusks, and vertebrates, even though chemical substances that have hormonal activity have been identified throughout the animal, plant, and fungal kingdoms. Even in vertebrates, there exists a class of hormones, the parahormones, designed to act locally. Because parahormones are made wherever they are needed, they tend to have a nearly ubiquitous distribution. Finally, many vertebrate growth factors are synthesized in multiple locations.
2. Blood for hormone distribution. First, blood is unique to vertebrates. The addition of hemolymph to the definition would permit arthropod hormones to be included in the definition, but those of plants and lower animals would still be omitted. Second, even in vertebrates, the parahormones diffuse through the extracellular fluid to reach their local targets. Other hormones are released by neurons and also have local effects. Finally, the classic definition would exclude ectohormones, hormones that traverse air or water to act between or among individuals. These hormones are particularly well developed in certain insect species and include pheromones (sexual attractants), gamones (inducers of sexual development), and allomones and kairomones (interspecies attractants).
3. A separate target organ. Some parahormones, once secreted, not only diffuse to surrounding cells but also stimulate the cells originally synthesizing them. This positive feedback is referred to as autocrine function, and it results in the synthesizing cell becoming its own target organ. Furthermore, bacteria make several regulatory molecules for internal use. These signal molecules, called alarmones, are usually modified nucleotides and are produced in response to a particular stress such as starvation or a vitamin deficiency.
Because of these limitations, a broader definition is used in this book: a hormone is a chemical, nonnutrient, intercellular messenger that is effective at micromolar concentrations or less. In other words, hormones are chemical substances that carry information between two or more cells. This definition includes all of the preceding examples except the alarmones. This exclusion is clearly the bias of mine, but the essence of endocrinology is the chemical coordination of bodily functions, and alarmones are used exclusively with single cells. However, other bacterial hormones that signal sporulation, competence (ability to take up exogenous DNA), conjugation, and other activities that are coordinated among individual bacteria are included. The restriction of hormones to chemical substances seems initially to be a logical one, even though species such as fireflies can use light to induce behavioral patterns in others. However, because the visual pigment rhodopsin and the G protein-coupled receptors (GPCRs) are homologous, one could also argue that, under certain circumstances, light is a hormone. Finally, metabolic pathways can be induced or repressed by substrate levels; indeed, substrate flow is an important regulator in many systems. Therefore nutrients are also excluded in the hormone definition. The inclusion of the concentration clause is used to eliminate other miscellaneous inducers; the one thing that sets hormones apart from other chemical regulators is their effectiveness at extremely low concentrations, usually in the nanomolar range or below. Plant hormones are unusual in that a few are required in larger amounts; it is for that reason that the micromolar limit is used.
The importance of endocrine regulation is apparent from the examination of the genomes of those organisms for which a complete sequence is available. For example, the genome of the nematode Caenorhabditis elegans contains about 20,000 genes. The single most abundant group, at 3.5% of the total, is the group of genes for GPCRs, which are receptors for hormones and other small molecules. The second most abundant group, at 2.6%, is the group of protein kinases, which are integral components of many signaling pathways. Finally, the third most abundant group, at 1.4%, is a transcription factor class that includes the nuclear receptors for steroids and other hydrophobic hormones. In metazoans, cellular communication and coordination are essential for successful development and survival, and their significance is reflected in the proportion of the genome allotted for endocrine functions.
The study of hormone action at the cellular and molecular level is called molecular endocrinology, which is the subject of this treatise. In particular, this book concentrates on the molecular mechanisms of hormone action and interaction. However, the topic of hormonal synergism and antagonism at the molecular level is better understood against a background knowledge of hormone action in the whole organism; for example, the progesterone inhibition of prolactin receptors and second messengers in the mammary gland is just an isolated fact unless one knows the general function of these hormones in the reproductive cycle. Therefore the function of this unit is to provide the reader, in general, and the novice, in particular, with sufficient background information to appreciate the molecular interrelationships that are discussed in later units. It is obvious that a complete presentation of general endocrinology cannot be accomplished in only three chapters: the coverage is specifically oriented and just sufficient to prepare the reader for the remainder of the book. However, it is hoped that the reader will become interested enough to consult any of the excellent and far more comprehensive texts listed in the General References section at the end of this chapter.
The rest of this chapter is concerned with identifying the basic characteristics of hormones and their regulation, and it concludes with an illustrative example, the hormonal control of calcium metabolism. Then, in Chapter 2, the other classical endocrine systems are examined. Finally, Chapter 3 briefly covers non-classical and nonvertebrate hormones, such as growth factors, parahormones, and the hormones of plants and insects.
Hormone-Target Relationships
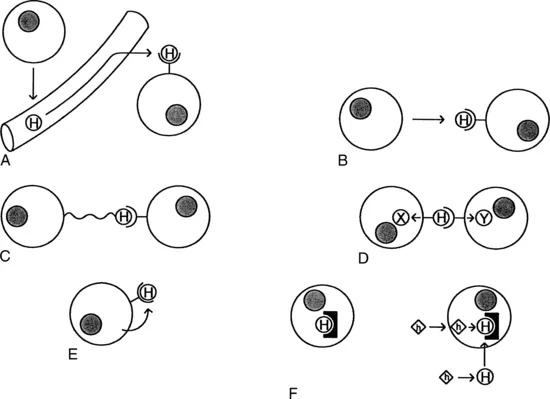
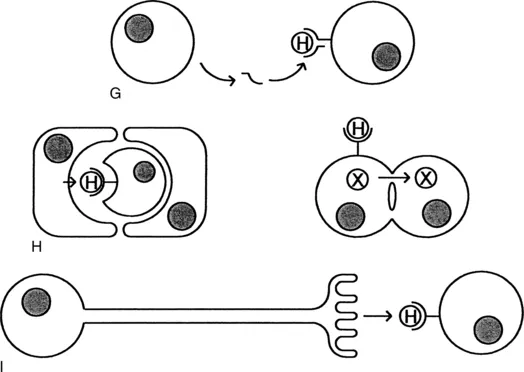
As noted previously, the classic endocrine system involves a hormone being made in one part of the body and reaching its target in another part of the body through the bloodstream (Fig. 1-1, A). However, there are many other types of interactions that can occur. In a paracrine system, the hormone remains in the tissue, where it reaches nearby cells by diffusion (Fig. 1-1, B). The juxtacrine system represents another mechanism for limiting the diffusion of hormones. In this case, the hormone is synthesized as a membrane-bound precursor. Although this precursor is usually cleaved to yield a soluble peptide, it may also remain attached to the plasma membrane, where it retains its biological activity. Therefore its effects are limited to the length of its tether (Fig. 1-1, C). Hormones that may act in this fashion include the epidermal growth factor, transforming growth factor α, tumor necrosis factor a, colony-stimulating factor 1, and the Kit ligand (see Chapter 3). In some cases, the intracellular domain of the hormone anchor is coupled to second messengers so that receptor engagement generates signals in both cells (Fig. 1-1, D). The ephrins are examples of this bilateral or reverse signaling. Finally, juxtacrine signaling may also include hormones whose diffusion is limited by the fact that they are tightly bound to the extracellular matrix.
The hormone may even influence the c...