![]()
1
Arms Races
As well as being the causative organisms of major human and animal diseases, parasites often serve as elegant models for the study of fundamental biological phenomena.
J. D. SMYTH (1994)
The Leitmotif of Life
The study of symbioses nowadays is always undertaken from a modern evolutionary perspective, that of Darwinism as it has been remodeled throughout the twentieth century and finally illuminated by discoveries in molecular biology. It is therefore important to recall that the significant thing in evolution is the reproductive success of individuals (or of populations), often termed fitness. If, for example, a pair of tits that lay eggs in a “clean” nest rear on average ten nestlings to fledging, whereas a pair that lay their eggs in a flea-ridden nest can raise only five offspring, one can say that parasitism by the fleas has lowered the reproductive success, or fitness, by half. (The real impact would be still greater if the offspring of the infested nest are of lower “quality” and therefore tend to survive for shorter periods.) In these terms, an advantage or benefit is thus augmented reproductive success, while a disadvantage or a cost is diminished reproductive success. Natural selection is the common thread that knits together all of evolution.1
To transmit their genes to the next generation appears as the leitmotif of all species that have existed on earth since the origin of life. When Charles Darwin enunciated the key principles of this process, he caused a scandal for several reasons. The main one was that without doubt he implicitly based all evolution (and right at the outset the origin of humans was at issue) on processes deprived of any moral or ethical content. People knew that wolves ate sheep and lions ate gazelles, but what could pass for a subsidiary clause of the punishment of Adam and Eve suddenly became, because of Darwin, a central mechanism of life. What am I saying? The central mechanism of all life!
Nearly a century and a half later, despite many discoveries, the principle of natural selection has never been cast into doubt. As for symbioses, they offer an excellent opportunity to discover and understand the workings of natural selection. If one imagines a caricature of a population of hosts confronted with a population of parasites, the hosts that are best at transmitting their genes to the next generation are those that defend themselves best against the parasites. Similarly, the parasites that transmit their genes best to the next generation are those that exploit their hosts best. We will see later that reality is not so simple but that this is a good description of the basic operation of natural selection. It implies reciprocal selective pressures. In a sense, the hosts select for the best parasites and the parasites select for the best hosts.
Encounter and Compatibility
We often compare predator–prey systems with those of parasites and hosts. Similarities do exist: in both systems, the interaction occurs only if there is an encounter. Examples are the encounter between a mouse and a cat in a predator–prey system and the encounter between an infective stage of a parasite and a mouse in a host–parasite system. But there follows a major difference between the two types of systems, as shown in figure 1.1.
In the predator–prey system, there is no post- encounter interaction. The mouse is eaten as soon as it is captured. What remains in the stomach and then the intestine of the cat during the brief period of digestion is no longer the mouse (and still less, the information that the DNA of the mouse carries) but common molecules from the cells that had made up the mouse.
By contrast, we can say that in the host–parasite system, the real action of the interaction actually begins after the encounter: either the mouse is able to destroy the infective stage of the parasite or the parasite is able to survive in the mouse. If the parasite survives, it manages to install itself in the right microhabitat and reproduces. This state can last for weeks or months, even for years. This second phase in the host–parasite relationship, after the encounter, is that of compatibility. We say there is a lasting, intimate interaction between the two partners (Combes 2001).
It is this lasting interaction that allows the hybridization and exchange of information that I discussed earlier.
Fig. 1.1. The fate of genetic information in a predator–prey system (A) and a host–parasite system (B). Only in the latter are the two sets of genetic information conserved side by side. This is a persistent, intimate interaction. Inf., information.
It is easy to see that from an evolutionary point of view it is in the best interest of the parasite to possess adaptations that allow it (1) to encounter its host and (2) to survive in the host if the encounter has occurred. Conversely, it is in the best interest of the host to have adaptations that allow it (1) to avoid encountering the parasite and (2) to get rid of the parasite if, despite any efforts to avoid an encounter, one has taken place. From these considerations arises an “arms race,” an expression that evokes the reciprocal selective pressures that the parasite species and the host species exert on one another’s evolution over long periods of time, even millions of years.
Let us imagine a host–parasite system at any time during its evolution and suppose that the parasite population is genetically diverse, so that certain individuals in particular infective stages behave in a genetically determined way that gives them a higher probability of encountering a host individual. Clearly, parasites that possess this behavioral trait will be positively selected, and their genes will tend to increase in frequency from generation to generation in the parasite population. For example, the behavior in question might be a positive response to a stimulus (odor, vibration, etc.) emanating from the host.
Now let us consider the host and imagine that certain individuals behave in a genetically determined way that reduces the probability that they will encounter an individual of the infective stage of the parasite. If the parasite is even slightly pathogenic, these host individuals that have avoided the parasite will be healthier than infected individuals. On average, they will be more successful reproductively, and their genes will tend to increase in frequency in the host population. The behavior that helps them avoid the parasite might be, for instance, fleeing to a habitat that the infective parasite stage cannot reach.
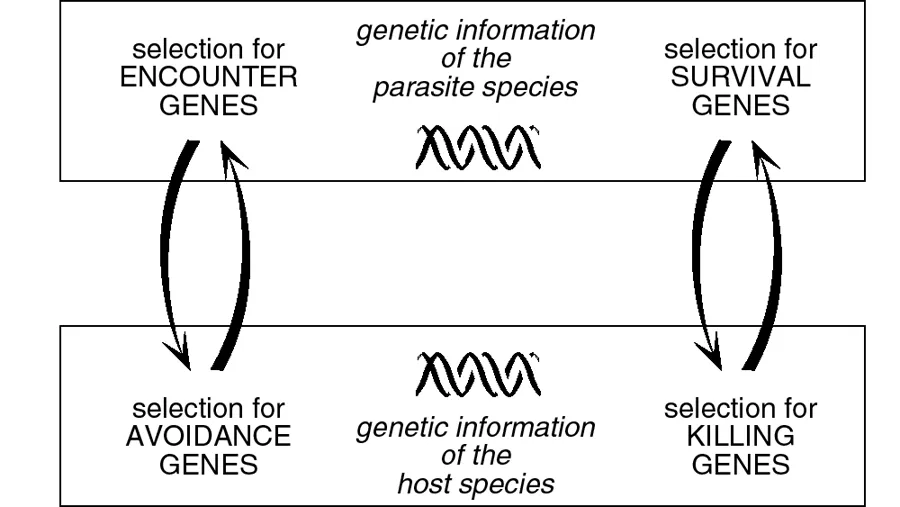
We see that these two selective mechanisms, one operating in the parasite population, the other in the host population, have all the features of an arms race. The more successful the parasites are in finding hosts, the more intense will be selection on the hosts, and the more beneficial to the host will be any adaptations that help it to avoid the parasite. The two species have thus engaged in an endless process. To the extent that the host becomes more effective at avoiding the parasite, the parasite survives only if it has sufficient genetic diversity that natural selection can produce better means of encountering the host. In turn, of course, selection in the host replies by producing new ways to avoid the parasite. In other words, selection for “encounter genes” in the parasite genome generates selection for “avoidance genes” in the genome of the host, and vice versa (fig. 1.2, left).
If we were dealing with a predator–prey system, things would stop here. The process I have just described can easily be applied to the relationship between cats and mice or between lions and gazelles. The cats and the lions have genes that help them encounter the mice and the gazelles, respectively. The mice and gazelles, in turn, have genes that help them avoid cats and lions. But in a host–parasite system, the story does not stop with the encounter.
Fig. 1.2. The two arms races in a host–parasite system: genes to facilitate encounters vs. genes for avoidance (left), and genes for survival vs. genes for efficient killing (right).
If an encounter has occurred, the host still has a chance to get rid of the parasite. The host possesses the astonishing property of being able to transform itself from the game to the hunter! All species (not only vertebrates but even the most primitive invertebrates) are able to recognize as foreign (that is, not-self) all molecules or ensembles of molecules that did not arise within them and to deploy against these intruders a battery of weapons. In the most highly evolved species, these weapons are extraordinarily elaborate, involving several sorts of cells (those that produce antibodies, those that are cytotoxic, etc.) and circulating molecules that are equally diverse (antibodies, cytokines, etc.).
The immune system mechanisms exert formidable selective pressure against pathogenic agents. The only parasite individuals that transmit their genes to the next generation are those whose traits allow them to survive in spite of the hostile milieu created by the host. Obviously, the selection of parasites able to survive maintains pressure on the host, so that natural selection allows host individuals to survive who have new weapons. All the ingredients are in place for a second arms race. The better the host can struggle against the pathogens, the more the pathogens are forced to adapt to the host’s armaments. Conversely, the better the parasite is able to cope with the host’s armaments, the more strongly natural selection forces the host to acquire new ones. The selection in the genome of the parasite for ability to survive in the host entrains selection in the host genome for ability to kill the parasite, and vice versa (fig. 1.2, right).
How the Arms Races Work
The two arms races I have just described can be represented by two filters—one for encounter, the other for compatibility. These two filters can be drawn as diaphragms (fig. 1.3). Natural selection operates in the parasite genome to open the two filters, and it operates in the host genome to close the two filters. What do these filters represent, from a genetic point of view?
These are hybridized phenotypes (Combes 2001), because their status at any moment depends not just on the genes of the parasite or on the genes of the host but on the genes of both individuals. These hybridized phenotypes obey the following rules.
1. The degree to which the filters are open determines not only the possibility or impossibility that parasitism will occur but also the abundance of the parasites if it does. This means there is very little chance that the degree of opening will remain constant. On the contrary, encounter and compatibility are both strongly influenced not only by eventual new mutations but also by environmental factors. For instance, a rainy year can favor completion of the life cycle of the large liver fluke because pastures are more humid (thus transmission is more frequent). Or an increase in the density of sheep can entrain a nutritional deficit and a consequent lowering of immune defenses (thus compatibility increases).
Fig. 1.3. Encounter and compatibility in host–parasite systems, represented by two diaphragms. Selection in the parasite genome tends to open the passage, whereas selection in the host genome tends to close it. The filters are hybridized phenotypes, undergoing opposing selection pressures in the parasite and host.
2. One closed filter alone suffices to keep the host from being parasitized. This fact confirms that hosts possess two sequential lines of defense and that, depending on the specifics of each particular case, selection can act more strongly on behavior that keeps the host from encountering the parasite or on immunity that protects the host after it has been parasitized. These alternatives Michael Hochberg (1997) has characterized by the phrase “hide or fight?”
3. The two arms races—of encounter and compatibility—are not independent in the sense that the adaptive responses are interchangeable. For example, to an increase in the frequency of encounters generated by natural selection acting on the parasite genome, the host can respond by increasing the efficiency of its immune system. The initial degree of parasitism would then be maintained.
As I have just described, host–parasite systems differ from predator–prey systems in one crucial detail. When a cat chases a mouse, both individuals run, but their goals are not the same. The cat runs in order to get a meal (after all, the cat will still be able to survive if it does not catch this particular mouse). The mouse, however, is running for its life (Dawkins and Krebs 1979). When an individual of an infective stage of a parasite seeks to infect a host individual, they both run (at least symbolically), but in this case it is the parasite that is running for its life, because it will die if it does not quickly find a suitable host. The host runs for its life only in particular cases; in general, it runs in order to be healthier.
This detail allows us to understand why there always are mice and parasites. The mice possess enough adaptations (to detect predators, to run quickly, to hide in inaccessible places, etc.) that a sufficient fraction of them escape cats. For their part, parasites possess enough adaptations both to encounter hosts (invisible infective stages, insertion in the food chain, use of vectors, etc.) and to survive even highly developed defense mechanisms (molecular mimicry, antigenic variation, immunosuppressive ability, etc.). Of course many infective-stage individuals die because they do not find a host, and many of those that do find hosts are killed after they infect them. Here also, however, enough parasites escape the slaughter, and the symbiosis persists from generation to generation.
Arms races also exist in mutualistic systems, even in obligatory mutualisms in which each of the two partners cannot survive without the other. Each partner (I return to this debate in chap. 5) remains fundamentally self-interested, and conflicts can linger for a very long time. Thus there are still conflicts between the mitochondrial genomes and nuclear genomes of eukaryote cells, even though this association is one of the oldest known. The only difference is that conflicts between mutualists are less severe (and thus less visible) then those between true parasites and hosts.
The health sciences today know ways of influencing just how open these filters are. Closing the encounter filter is achieved by education and hygiene measures that lead to the avoidance of infective stages and contagion. As for closing the compatibility filter, this is achieved in two ways: (1) by therapy (for example, by antibiotics, either synthetic or natural) that suddenly renders the ...