
- 242 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
A Transformation Theory of Aesthetics
About this book
This book develops a theory of picture perception and aesthetic response, arguing that images can generate in us a complex pattern of mental changes, or transformations. It is essential reading to those seriously involved in linking the arts and cognitive sciences.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
PhilosophySubtopic
Aesthetics in PhilosophyChapter One

THE BIOLOGICAL AND PROCEDURAL BASES OF NEUROPSYCHOLOGY
1.1 INTRODUCTION
I have proposed that neuropsychology can illuminate certain mental competences that have a bearing on the meaning of art. This opening chapter clarifies the physical bases of the brain relevant to neuropsychology and reviews the more essential procedures it has developed to reach its findings. I shall first make a few opening remarks concerning the definition and implications of neuropsychology.
It has long been established that aspects of cognition which would be difficult to study in the healthy individual under laboratory conditions may break down in a circumscribed manner as a result of brain injury, surgery or disease. The core of neuropsychological research proceeds from a study of the clinical data provided by such cases. Even so, because neuropsychology is concerned in general with how mental life and human behaviour is related to the structure of the brain, the study of the healthy individual is not excluded from its research, as Dimond indicates.
We are certainly not going to exclude knowledge merely because it comes from somewhere else such as the study of animals with experimental lesions of the brain or the study of the normal person who has his brain in one piece.(Dimond 1978: 4)
Taken collectively, neuropsychology’s findings clearly indicate that the human brain possesses qualitatively distinct modes of cognitive processing that can be cerebrally localized. It is for this reason that it is often seen as complementing faculty psychology. In recent years, however, perhaps the most significant and certainly the most dramatic insights regarding cerebral localization of function have come from experiments conducted on commissurotomy or ‘split-brain’ subjects.1 This branch of neuropsychology is termed lateralization research because of its assumption that certain higher level mental competences are lateralized to either the left or right cerebral hemispheres. Nonetheless, in keeping with the general description of neuropsychology, lateralization research includes the study of the extent to which competences are lateralized in the cerebrally intact human brain.
Cerebral lateralization of function should not be taken to mean, however, that cerebrally ‘normal’ individuals act out dualistically motivated lives where the left hand has no knowledge of what the right is doing, as it were. Such cases have been known to occur, but only in the commissurotomized and then only rarely.2 As Roger Sperry, the leading investigator in the field, indicates, those who find insights gained from lateralization research should be wary, because:
In some cases the conclusions, along with the growing wave of semi-popular extrapolations and speculations, concerning the ‘left brain’ versus ‘right brain’ functions, call for a word of caution … in the normal state, the two hemispheres appear to work closely together as a unit.(Sperry 1982: 1225)
Many have heeded Sperry’s advice, ranging from those involved in direct experimentation in the field (Bryden 1982, for example), to those concerned with its implications for other fields (art education, for example; see Youngblood, 1979), or art in general (Schweiger 1985, for example).3
Nonetheless, inherent in Sperry’s caution is the implication that successful cognition is contingent upon a co-operation of the hemispheres’ modes of functioning. From this implication I shall argue in later chapters that when the hemispheres are presented with certain forms of pictorial information, their associated modes of processing do not necessarily work in harmony. But these are early times in this book and the purpose of this introductory chapter is not to overly speculate but to clarify the essential bases from which neuropsychology operates.
1.2 THE CEREBRAL CORTEX
Higher mental functioning in humans is associated with the cerebral cortex, a deeply fissured mantle of the brain divided into two visually like areas known as the left and right cerebral hemispheres. Each hemisphere is subdivided into four major areas known as the occipital, temporal, parietal, and frontal lobes.4 The mirror-image nature of the cortex reflects the left-right symmetry of the body. However, although the control of basic body movement and sensation is symmetrically distributed between the hemispheres, the control mechanisms operate in reverse, as it were. In general, the left hemisphere controls the right side of the body, the right hemisphere the left, thereby conceptually establishing a cross-fashion or ‘X’ functional mode. This is physically substantiated by the fact that the left hemisphere’s neural connections receive and generate signals primarily to the right side of the body and vice versa.
As a result of this X functional mode, if inputted information is confined to one side of the body it is initially primarily contralaterally registered by the cortex. For example, tactile information experienced by the right hand is primarily registered by the left hemisphere, whereas information from the left hand is registered by the right hemisphere. The notable exception is olfactory information which is ipsilaterally registered. That is, the left nostril transmits information to the left hemisphere, the right nostril transmits information to the right hemisphere.
The same X functional mode applies to the generation of basic body movement, that is, the brain’s basic capacity to control motor skills. Motor skills for the movement in the hand or foot, for example, primarily emanate from the hemisphere located on the opposite side of the body. The exception is the movement of facial muscles which are controlled by both hemispheres.
It should be emphasized, however, that the way in which the brain controls basic body movement and sensation should not be confused with the way in which it processes and subsequently responds to higher order information, in which case it displays a marked asymmetry of function. In right-handers, for example, it is almost exclusively the case that the hemisphere controlling the dominant hand is also the one controlling speech, that is, the left hemisphere. There is, however, a great deal of accumulated evidence which argues that, in the case of the right-hander drawing a picture, for example, the guiding spatially oriented knowledge emanates from the right hemisphere.
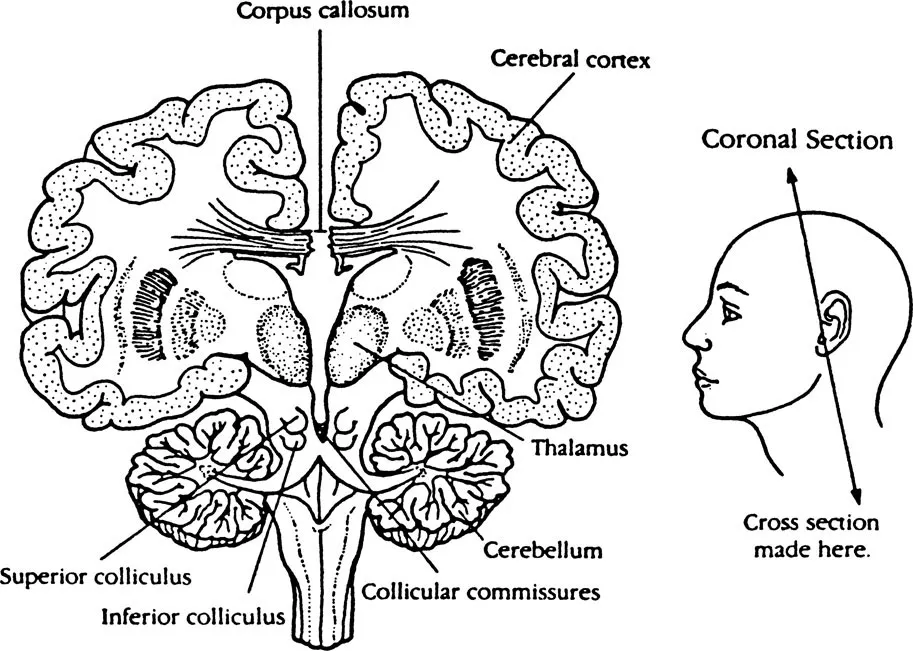
Figure 1 Extent of the separation of the brain following forebrain commissurotomy
From this it follows as a general rule that the hemisphere which generates the basic body movement relevant to a particular task is not necessarily the one most capable of its cognition.
At this point, however, Sperry’s indication that the hemispheres normally function as a unit should be emphasized. This is because they are linked to the spinal column and peripheral nerves through a common brain stem and because, above all, they are linked by the corpus callosum, a large band of nerve fibres embedded between the hemispheres. These connections are collectively known as the commissures. Most of the evidence indicates, however, that it is primarily the corpus callosum that enables higher order information to pass in some form between the hemispheres. As a result, even if information is initially exclusively registered and processed by one hemisphere, it can be transferred to its partner hemisphere (exactly what transformational laws this transference might involve is still unknown).
The exception is found in the commissurotomized because, with the corpus callosum disconnected, information can no longer be directly exchanged between the hemispheres (Figure 1). This disconnection means that the hemispheres become functionally isolated from each other thereby enabling their respective cognitive characteristics to be individually studied.5 This study requires complex procedures to ensure that information inputted to the cortex is registered exclusively by the hemisphere under investigation, that is, the information must be unilaterally registered.
At this point, then, something should be said about the procedures developed by neuropsychology to confine information to one hemisphere. Some of the procedures used to study the brain-damaged are also used to study the normal subject.
1.3 TACHISTOSCOPIC PRESENTATION
The unilateral registration of visual information presents the researcher with a particularly complex problem. This is because in both the normal and brain-damaged subject each retina can relay information to the cortex so that it is both contralaterally and ipsilaterally registered.6 That is, the left half of each retina, which scans the right half of the visual field, sends impulses to the left hemisphere; impulses from the left half of the visual field are transmitted by the right half of each retina to the right hemisphere. What this amounts to for experimental purposes is that if the subject fixates on a central point in front, the right hemisphere exclusively registers information presented to the left visual field (LVF) whereas the left hemisphere exclusively registers information presented to the right visual field (RVF). However, to ensure correct hemispherical registration the speed at which information is presented to either visual field has to be faster than the saccadic eye movements of the subject. This is because if their focus strays even momentarily from the central point to either visual field, the information will be bilaterally registered, that is, registered by both hemispheres.
The need to present visual information at speed necessitated the development of the tachistoscope, a device which flashes information to either side of a screen with a marked central point. In this way, when focusing on this point, information flashed to either side falls exclusively within the subject’s respective visual fields and is therefore unilaterally registered as required (Figure 2).
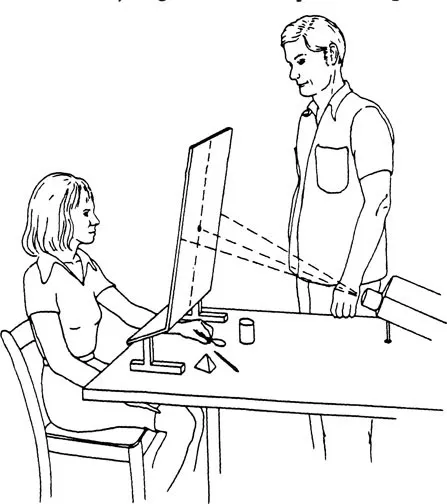
Figure 2 The basic testing arrangement used to lateralize visual and tactile information and allow tactile responses
While tachistoscopic presentation to the commissurotomized solved the problem of limiting information to either hemisphere, the speed at which it had to be presented limited experimentation; that is, the subject could not be expected to detail a response to information viewed for only a split second. This limitation was overcome, however, when Zaidel (1975) developed a new method of restricting visual information to one hemisphere. It utilizes a device described as the Z contact lens occluder (the Z lens) which permits subjects to freely move their eyes when examining information and at the same time ensures that it is registered by only one hemisphere.
The Z lens therefore makes it possible to view information for as long as necessary to an experiment yet also allows it to be exclusively registered by the hemisphere under investigation. The use of the Z lens has led to some of the most important lateralization research studies centred on establishing the right hemisphere’s level of consciousness, as we shall see in the next chapter.
1.4 CROSS-MODAL REFERENCING7
Early tachistoscopic presentation involving commissurotomy subjects quickly established that information flashed to the LVF (right-hemisphere registered) went verbally unacknowledged. This was predictable because it was known that, although the right hemisphere possesses some capacity to understand language, in the majority of cases the speech centres are left-hemisphere located. Researchers therefore had to find extra-verbal methods of eliciting from the subject the extent to which information had been registered and comprehended by the right hemisphere.
One of the commonest methods was to flash an image of a well-known item – a spoon, for example – and then to ask the subject to select, using the left hand, a real spoon from an array of items hidden from view.8 In most instances, the subject correctly identified the item by feeling its shape and contours (Figure 2). This clearly indicated that, although the item’s image had gone verbally unacknowledged, it had, in fact, been right-hemisphere registered and comprehended. Nonetheless, unless the item was shown to the left speech hemisphere, it continued to remain verbally unacknowledged by the right.
Other methods of eliciting the extent to which tachistoscopically presented information had been right-hemisphere registered and comprehended were developed by researchers, leading to some intriguing results.
1.5 CROSS-CUING
As lateralization research developed, certain inconsistencies in the findings began to occur with increasing frequency. Subjects initially unable to verbally identify items hidden from view but held in the left hand began to name some items. Interpretations of these results ranged from the suggestion that over time the right hemisphere had developed a speech capacity to the suggestion that information was being transmitted between the hemispheres via the lesser communication pathways such as the lower brain stem.
Gazzaniga and Hillyard (1971) offered a simpler explanation for the inconsistency which they termed cross-cuing. This refers to the ...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Original Copyright Page
- Dedication
- Table of Contents
- List of figures
- Foreword
- Introduction
- 1 The Biological and Procedural Bases of Neuropsychology
- 2 The Experimental Evidence
- 3 The Possible Origins of Cerebral Specialization
- 4 The Right-Hemisphere Cognitive Paradigm
- 5 Children’s Drawing
- 6 The Languages of Art
- 7 A Transformational Theory of Aesthetics
- 8 Art and Aesthetic Experience
- 9 The Transformational Planes
- Appendix: Psychoanalysis and Art
- Notes
- Bibliography
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access A Transformation Theory of Aesthetics by Michael Stephan in PDF and/or ePUB format, as well as other popular books in Philosophy & Aesthetics in Philosophy. We have over 1.5 million books available in our catalogue for you to explore.