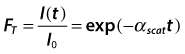Ocular Disease—a newly introduced companion volume to the classic Adler's Physiology of the Eye—correlates basic science and clinical management to describe the how and why of eye disease processes and the related best management protocols. Editors Leonard A. Levin and Daniel M. Albert—two of the world's leading ophthalmic clinician-scientists—have recruited as contributors the most expert and experienced authorities available in each of the major areas of ophthalmic disease specific to ophthalmology: retina, cornea, cataract, glaucoma, uveitis, and more. The concise chapter structure features liberal use of color—with 330 full-color line artworks, call-out boxes, summaries, and schematics for easy navigation and understanding. This comprehensive resource provides you with a better and more practical understanding of the science behind eye disease and its relation to treatment.- Covers all areas of disease in ophthalmology including retina, cornea, cataract, glaucoma, and uveitis for the comprehensive information you need for managing clinical cases.- Presents a unique and pragmatic blend of necessary basic science and clinical application to serve as a clinical guide to understanding the cause and rational management of ocular disease.- Features 330 full-color line artworks that translate difficult concepts and discussions into concise schematics for improved understanding and comprehension.- Provides the expert advice of internationally recognized editors with over 40 years of experience together with a group of world class contributors in basic science and clinical ophthalmology.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Loss or reduction in corneal transparency occurs from a variety of causes, including edema resulting from diseases such as Fuchs’ dystrophy and bullous keratopathy, scarring resulting from wound healing, haze following photorefractive keratectomy, and certain metabolic diseases such as corneal macular dystrophy. The intent of this chapter is to review the present understanding of mechanisms or structural alterations that cause loss of corneal transparency. Transparency loss resulting from edema, scarring, and photorefractive keratectomy will be emphasized.
Understanding the mechanisms of transparency loss requires understanding the structural bases of corneal transparency itself. Because the cornea does not absorb light in the visible portion of the electromagnetic spectrum, its transparency is the result of minimal light scattering.1,2 Visible light is an electromagnetic wave with wavelengths between 400 and 700 nm. Light scattering results when an incident light wave encounters fluctuations in the refractive index of a material. These fluctuations cause some of the light to be redirected from the incident direction, thus reducing the irradiance in the forward direction. The transmissivity, FT, is defined as:
(1)
where I(t) is the irradiance of the light transmitted through a scattering material of thickness t (e.g., the cornea), I0 is the irradiance of the incident light, and αscat is the extinction coefficient due to scattering.3,4 As will be shown in the remainder of this chapter, the quantity αscat provides significant information on the nature of the structural features responsible for the scattering.
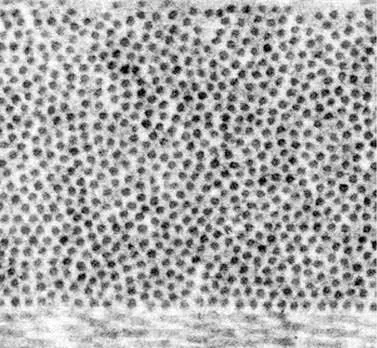
Collagen fibrils, which lie parallel to one another within the lamellae of the corneal stroma, have a somewhat larger refractive index than the optically homogeneous ground substance surrounding them. Thus they scatter light. In fact, because they are so numerous they would scatter approximately 60% of an incident beam of light having a wavelength of 500 nm if they were randomly arranged like gas molecules and therefore scattered independently of one another (i.e., FT would be 0.40).1,5 A normal cornea scatters only about 5% of 500 nm light1; thus transparency theories seek to explain why the scattering is so small (Box 1.1). The key is that destructive interference among the scattered fields, which arises because the fibrils possess a certain degree of spatial ordering about one another, reduces the scattering that would otherwise occur. Indeed, Maurice’s lattice theory of transparency postulated that the fibrils within the stromal lamellae are arranged in a perfect hexagonal lattice. Because their spacing (which is approximately 60 nm) is less than the wavelength of visible light, Bragg scattering cannot occur and such an arrangement leads to perfect transparency.5 Obviously the corneal stroma is not perfectly transparent. If it were, it could not be visualized in the slit-lamp microscope. Although scattering from keratocytes could be used to explain visibility in the slit lamp, all present evidence suggests that they are not a significant source of scattering in normal cornea except under the specialized condition of specular scattering that occurs in confocal images or in the slit lamp when the incident and viewing directions are configured to make equal angles with the surface normal.1,2,6,7 Additionally, transmission electron micrographs (TEM) of the normal stroma do not depict a perfect lattice arrangement (Figure 1.1). Thus, as described in the remainder of this section, investigators have built on the Maurice model by relaxing the condition of perfect crystalline order.
Box 1.1 Characteristics of light scattering in normal cornea
• The matrix of collagen fibrils is the major source of light scattering in normal cornea
• Keratocytes are not a significant source of scattering in normal cornea except under the specialized condition of specular scattering
• Measurements of how the total scattering cross-section depends on light wavelength can be used to distinguish between the various transparency theories
Figure 1.1 Transmission electron micrograph of the posterior region of a human cornea. The fibrils are shown in cross-section.
Scattering from an array of parallel cylindrical collagen fibrils is characterized by a quantity σt(λ), called the total scattering cross-section. It is equal to σ0t(λ)σtN(λ), where σ0t(λ) is the total scattering cross-section per unit length of an isolated fibril, σtN(λ) is the interference factor, and λ is the wavelength of light in the stroma.8 The total scattering cross-section per unit length of an isolated fibril, σ0t(λ), depends on the fourth power of fibril radius and the ratio of the fibril index of refraction to that of its surroundings and its wavelength dependence is inverse cubic (i.e., σ0t(λ) ~ 1/λ3).3,4 The interference factor, σtN(λ), is the subject of all modern transparency theories.5,9-12 These have been reviewed extensively elsewhere and will not be discussed in detail here.1,2,13 The value of the interference factor varies between zero (for Maurice’s perfect lattice theory) and one (for fibrils with random positions – the independent scattering result discussed above). In order to agree with experimental values of transmissivity, its value is about 0.1 at a wavelength of 500 nm (Box 1.2).
Box 1.2 Factors underlying corneal transparency
Corneal transparency is due to three major factors:1,2,13
• Individual fibrils are ineffective scatterers because of their small diameter and their refractive index is relatively close to the surrounding ground substance (the ratio is ~ 1.04)
• Destructive interference among the ...
Table of contents
Cover
Title Page
Front Matter
Copyright
List of Contributors
Dedication
Foreword
Preface
Table of Contents
SECTION 1: Cornea
SECTION 2: Dry eye
SECTION 3: Glaucoma
SECTION 4: Lens
SECTION 5: Neuro-Ophthalmology
SECTION 6: Oncology
SECTION 7: Other
SECTION 8: Pediatrics
SECTION 9: Retina
SECTION 10: Uveitis
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Ocular Disease: Mechanisms and Management E-Book by Leonard A Levin,Daniel M. Albert in PDF and/or ePUB format, as well as other popular books in Medicine & Opthalmology & Optometry. We have over 1.5 million books available in our catalogue for you to explore.