
- 160 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Basis of Progressive Evolution
About this book
In this incisive book, a distinguished geneticist has succeeded in relating the extraordinary biological discoveries of the last two decades to the basic questions about the origin and evolution of life on earth. The "molecular revolution" in biology--culminating in the discovery of the relations of the giant molecules of living matter--the operation of the genetic code, and the relational order in the biological world are all considered.
Originally published in 1969.
A UNC Press Enduring Edition -- UNC Press Enduring Editions use the latest in digital technology to make available again books from our distinguished backlist that were previously out of print. These editions are published unaltered from the original, and are presented in affordable paperback formats, bringing readers both historical and cultural value.
Originally published in 1969.
A UNC Press Enduring Edition -- UNC Press Enduring Editions use the latest in digital technology to make available again books from our distinguished backlist that were previously out of print. These editions are published unaltered from the original, and are presented in affordable paperback formats, bringing readers both historical and cultural value.
Tools to learn more effectively

Saving Books

Keyword Search

Annotating Text

Listen to it instead
Information
1: LEVELS OF ORGANIZATION AND THEIR EVOLUTION
The “revolution in biology” that has swept through the world of science during the past twenty years is beginning to affect our concept of the nature of the world and of life to a far greater degree than most scientists realize. Furthermore, I believe that it is destined to have even more profound effects upon our thinking during the next twenty years.
In the minds of most biologists and biochemists, the biological revolution has centered about the discovery that nucleic acids, particularly DNA, are the chemical basis of heredity and that the specificity of proteins, the “building blocks” of the body, is based upon the translation of a “genetic code” that consists of specific sequences of nucleotide bases. This is, of course, the kernel and fountainhead of the revolution. Nevertheless, its effects are spreading far beyond these particular relationships. In the present chapter, the breadth and depth of these effects will be probed, while the remaining chapters will discuss some of them in greater detail.
Life Depends upon Organization
Perhaps the most significant of these broader effects of our understanding of the genetic code is confirmation and reinforcement of a belief previously held by most biologists. The essence of life is not a particular substance or substances, but a particular kind of organization. In textbooks, this belief has been expressed by likening an organism to a candle flame. Just as in a quiet atmosphere, the flame keeps its shape in spite of the fact that new particles of wax are being continually fed into it while the remains of burned particles are rising in smoke, so the living cell keeps its shape in spite of the constant replacement by synthesis and degradation of the molecules that compose it. Life as organization is a concept that is also supported by our understanding of death. When a man, an animal, or a cell dies, it does not necessarily lose any substance in the act. Death is a loss of function rather than substance. It is based upon changed relationships between organs, tissues, or cells, or between the molecules of which the cell consists. More will be said about this topic at the end of the present chapter.
The Principle of Relational Order
The statement that life is both orderly and complex is a truism that by itself tells us little. Recent discoveries concerning the molecular structure of living matter have, however, revealed two facts about the nature of this order which are of the utmost significance. In the first place, the macromolecules of which living matter chiefly consists, the proteins and nucleic acids, owe their specificity to particular linear arrangements of the smaller molecular units that make them up. The order of living macromolecules is not that of orbits within orbits, as is that of the solar system as well as of the atoms composing it; nor is it the order of geometrical crystals. Rather, it is like the order of a printed page or a taped message, the symbols of which are like the letters of the alphabet or the dots and dashes of a telegraphic code. Secondly, the ordered sequences of protein molecules, which are the working units of living systems, are perpetuated not by any properties that are inherent in their own structure but by a complex relationship existing between this order and that found in the nucleic acids that are the vehicles of heredity. The molecules of nucleic acid are like a designer’s blueprints for the construction of a machine tool, a work bench, or the parts of a building, written in a telegraphic shorthand that consists of four symbols. Given the environment of the living cell or of certain ones of its components, particularly the enzymes and coenzymes necessary for translation, the order of units (purine and pyrimidine bases) in a molecule of nucleic acid specifies precisely the order of units (amino acids) in a molecule of protein. The two kinds of molecules bear a colinear relationship to each other. This colinear relationship is the basis of both heredity and functional order in living systems; its alteration through changes in the base sequences of nucleic acids, which we now recognize as the chemical basis of genetic mutation, provides the hereditary variability necessary for evolution.
Other relationships between macromolecules, as well as between ordered combinations of them, also play vital functional roles in living systems. They form a hierarchy of order with at least eight levels: macromolecule, macromolecular aggregate, organelle, cell, tissue, organ, system of organs, individual. At each level, precise relationships between equally complex structures of different kinds are necessary for ordered function. We are all aware of such relationships at the level of organs such as nerves and muscles and of organ systems such as the respiratory system that regulates our breathing and the circulatory system that distributes oxygen through our bodies. Functional relationships at the levels of tissue, cell, organelle, and macromolecular aggregate will be described below.
Recognition of the hierarchy of ordered structures that makes up the living organism leads us to a basic principle about the nature of life. This can be designated as the principle of relational order and defined as follows. In living organisms, the ordered arrangement of the basic parts or units of any compound structure is related to similar orders in other comparable structures of the same rank in the hierarchy, permitting the structures to cooperate in performing one or more specific functions. The functions that depend upon relational order can be as diverse as muscular contraction, the transmission of nervous stimuli, the manufacture of sugar by photosynthesis, the synthesis of the protein molecules that form the building blocks of the living cell, and the transmission of heredity. Consequently, relational order is of primary importance as a unifying principle for all of life.
Relational Order and the Activity of Living Systems
A basic feature of relational order is that it is not a series of static relationships, like those between the molecules of a chemical crystal or between the beams and rafters of a house. The ordered relationships among parts of all levels of the organismal hierarchy help organisms both to carry out certain chemical reactions and to bring about co-ordinated movements of parts at all levels. At the molecular level, in fact, many of the chemical reactions of metabolism are inextricably bound up with, and dependent upon, the orderly movements of macromolecules and small organelles, such as the protein fibrils found in muscle cells (Figure 1–3).
The precise relational order between the macromolecules that make up cellular organelles is also in many instances associated with ordered movements. The regular beatings of cilia that propel the cells of protozoa such as Paramecium through the water are made possible by a complex organized system of protein filaments. The cilia themselves have an elaborate, stereotyped structure of nine peripheral protein filaments that are contractile and two central filaments that probably conduct stimuli. Co-ordination is achieved by a complex network of filaments that connect cilia with each other. Even more remarkable are the regular movements of chromosomes and associated structures during mitotic cell division. Mitosis is, in fact, an ideal illustration of how orderly movements of organelles that have been made visible to us depend upon equally orderly, systematic movements of individual molecules about which we still can only make inferences. This is true in spite of the fact that the molecular activities accompanying mitosis are still very imperfectly known. The bridge between visible movement and molecular behavior is being built by three kinds of research activities: detailed motion pictures of cells and chromosomes in mitosis both under normal and abnormal conditions; electron micrographs of dividing cells, in both mitosis and meiosis; and biochemical studies of the orderly succession of different cellular environments or states that is essential for the course of mitosis.
Motion pictures, particularly those of Andrew Bajer, have shown that the separation of daughter chromosomes from each other depends upon the contraction of specific, recognizable protein fibrils. Furthermore, this contraction is like that of muscle fibrils in that it involves changes in the shape but not in the volume of the molecules concerned. Electron micrographs have revealed the geometric structure of these fibrils and have shown that other protein fibrils, much like them, exist in the cell before mitosis begins (Figure 1–1). In fact, the orientation of these fibrils, known as microtubules, may in some instances be a predetermining factor that dictates that a cell will divide in a particular direction, long before the division itself begins.
Physiological and biochemical studies have shown that the orderly movements of chromosomes and other structures during mitosis are correlated with an orderly succession of different cellular environments. Oxygen consumption increases just before mitosis begins and drops while the process is going on. Soluble sulfhydryl (SH) groups show a similar rise and decline. Mitochondria, the energy supplying organelles or “engines” of the cell, execute ordered movements correlated with stages of the mitotic cycle. Regular changes in the viscosity of the living matter also accompany mitosis. While the meaning of all of these changes is not yet clear, we can hardly doubt that they have a direct connection with the visible movements of organelles that make cell division possible.
Muscular Activity and the Hierarchy of Order
The hierarchy of order in structure and function is well illustrated by the activity of muscle. Starting with the
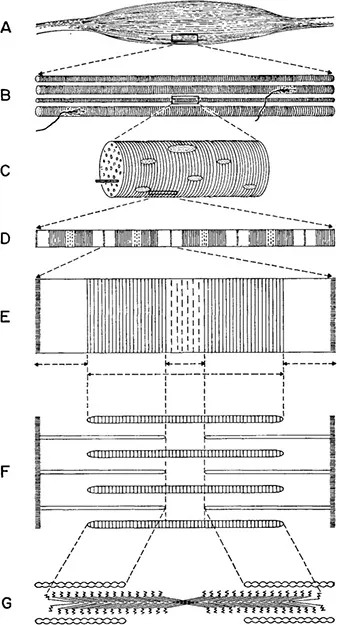
Figure 1–3. The hierarchy of complexity in muscle tissue. A, a single muscle, similar to that shown in the hand (Fig. 1-2), magnification x2. B, a group of fibers, with nerve endings attached, x100. C, portion of a single fiber, x1000. D, portion of a single fibril, showing striations, x10,000. E, the indicated portion of D, x43,500. F, the same, showing relative positions of the thick and thin filaments, which are, respectively, groups of myosin and actin molecules. G, diagram showing the arrangement of individual myosin molecules in a thick filament and portions of the rope-like structure that consists of actin molecules wound together. The “heads” of the myosin molecules are shown as zigzag lines pointing toward the actin filaments. Magnification x130,500. If the entire muscle shown in A were magnified to the same scale as the molecules shown in G, it would be more than 5 miles long!
From H. E. Huxley in Scientific American 199(5), 1958, and 213(6), 1965. Reproduced by permission.
organism as a whole, we all recognize that the eyes, ears, and other sense organs are continually sending messages to the brain, where they are sorted out. Impulses then go to various muscles, which respond in ordered ways. At this level, we see the brain as the ultimate center of integration and order, while all of the bodily organs can be regarded as merely tools for carrying out its directives. We can go one step down in the hierarchy by focusing our attention upon a single organ, for instance the hand. If we dissect away the skin from a human hand, we see a labyrinth of muscles and tendons (Figure 1–2). Each muscle is supplied by a nerve or nerve branch that comes from the brain. In addition, the skin is well supplied with nerve endings, which give us a sense of touch. The coordinated action of nerves, brain, and muscles enables the hand to do anything from avoiding a flame to feeling the nature of an object or grasping a rope, a bar, or a tree branch. For each of these actions, however, localized co-ordination is required between sense organs, nerves, a part of the brain, and the muscles of the hand.
Continuing down the hierarchy, we see a new level of co-ordination within the muscle itself. Isolated muscles, kept at the right temperature and bathed in saline solution, can contract in response to artificial stimuli that simulate nervous impulses. If we dissect a muscle, we find that the contraction of the organ as a whole actually consists of the co-ordinated activity of hundreds of muscle fibers, each of which is simultaneously stimulated by an individual nerve cell or neuron. Before we can understand what happens at the molecular level, we must focus our attention upon individual muscle fibers.
Of the numerous scientists who have supplied information bearing upon the relationship of this ordered structure to muscular contraction, one of the leaders is Professor H. E. Huxley of University College, London, whose account of it is followed in the present description.
In its superficial appearance, each fiber is like a tube surrounded by a protein membrane. When the muscle is inactive, a small difference in electrical potential exists between the outer and inner surface of this membrane. If this difference is temporarily eliminated, muscular contraction results. The fiber itself is a single cell containing many nuclei. It also contains mitochondria, tiny organelles that in most cells supply energy through the breakdown of sugar and other carbohydrates. The ability of muscles to contract, however, depends upon the presence in the fiber cells of many thread-like fibrils to thin that they are barely visible under the light microscope. Under the electron microscope, each fibril is seen to contain thousands of filaments. The latter are of two kinds in respect to thickness (Figure 1–3). Chemical analyses have shown that the thick filaments differ from the thin ones in respect to their protein composition: that of the thick filaments is myosin; and of the thin filaments, actin.
Biochemical studies, in which the individual molecules of myosin and actin have been separated from each other and put together again, have been combined with detailed observations under the electron microscope. This research has led to an ingenious and highly plausible hypothesis on the way in which myosin and actin molecules interact to cause the muscle to contract. When myosin molecules are isolated and photographed at very high magnifications under the electron microscope, they are found to consist of a globular head and a very long threadlike tail. The head has two functions. It can release chemical energy for the work of muscular contraction by carrying out the most widespread energy producing reaction in living systems: removing a phosphate group from a molecule of the ever-present compound, adenine triphosphate (ATP). It can also form a cross-link that binds the myosin molecule chemically to one of actin. The tails of adjacent myosin molecules stick together to form the thick filaments. The molecules of actin are bead-like but are joined together to form a twisted cable, which is the thin filament.
A limb muscle in the relaxed condition is striped or striated, the bands consisting of regions containing both thick and thin filaments that alternate with regions containing only thin filaments. At each end of the thick filament, myosin molecules are so arranged that their hands point endwards, and the middle part of the molecule contains only molecular tails. If, therefore, the changed cellular environment produced by the nervous stimulus alters in some way the binding properties of actin-myosin crosslinks, these can cause the actin filaments to slide over the thick filaments of myosin and thereby bring the muscle to its contracted state. The action is likened by Dr. Huxley to that of a ratchet, which in a typewriter turns the cylinder to which the paper is attached.
From these studies we have learned that the inside of a muscle-fiber cell is completely different from the foam- or jelly-like structure that, according to the biologists of half a century ago, was the nature of the “protoplasm” contained in a cell. Although the muscle cell is an extreme example of molecular complexity, other cells approach it to varying degrees. As will be explained in the next chapter, enzyme molecules can be likened to complex tools, the parts of which are precisely adjusted so that they can interact to perform very specific chemical functions and in many instances can be “turned on” or “turned off” by signals or switches that other molecules provide. Furthermore, most enzymes are not irregularly distributed through the cell but are located in precise positions on organelles such as mitochondria and plastids, on membranes known as the endoplasmic reticulum, or are embedded in the membrane that bounds the cell. The inside of a cell is not the “bubble bath” that biology students of an earlier generation were led to expect. Rather, it is like an intricate system of carefully placed and adjusted molecular machinery or like a factory room studded with benches bearing various kinds of tools and machines.
The Energy Supply for Living Systems
Biologists are well aware of the fact that the molecular machinery requires energy for its activity, and have probed deeply both into the ultimate source of this energy and the form in which it is used by the working molecules. Animals obtain energy by breaking down the complex organic molecules of their food; plants obtain it from sunlight. But whatever is the source of the energy, its transfer and immediate availability for work is accomplished chiefly by a single widespread kind of chemical reaction, the reversible transformation of nucleotide triphosphates, chiefly adenine triphosphate (ATP), into diphosphates. We can realize most graphically the difference between the way man runs his machines and the way in which the molecular machinery operates by making two analogies. The first one is between a generator or transformer and a mitochondrion, the kind of organelle in which sugars are broken down to release energy. These are all devices for transforming one kind of energy into another. The second is between a small electric motor and a molecule of ATP plus that portion of an an enzyme or coenzyme which serves to break the phosphate bond so as to release its energy.
Mitochondria contain thousands of molecules of the related and interchangeable compounds, adenosine diphosphate (ADP) and adenosine triphosphate (ATP), the latter of which is the universal energy carrier of living systems. These molecules may be likened to tiny motors. The conversion of ADP to ATP is comparable to “winding up” the motor, since it requires an input of energy; while the reverse reaction, ATP—ADP, is usually one by which energy is released for the work of the body, whether it be the synthesis of its molecular architecture or the ordered movements that keep it going.
A man-made factory usually has only one or a few generators or transformers and a limited number of motors. Since apparatus of this kind is difficult and expensive to design, build, and run, efficiency in the factory is greatest if energy is delivered in the form of high voltage elect...
Table of contents
- Cover Page
- THE BASIS OF PROGRESSIVE EVOLUTION
- Copyright Page
- PREFACE
- CONTENTS
- 1: LEVELS OF ORGANIZATION AND THEIR EVOLUTION
- 2: THE GENETIC CODE AND ITS EVOLUTIONARY IMPLICATIONS
- 3: DEVELOPMENTAL INTEGRATION AND EVOLUTIONARY PROGRESS
- 4: THE GENERAL NATURE OF EVOLUTION
- 5: THE EVOLUTION OF SOCIAL ORGANIZATION
- SUGGESTED ADDITIONAL READING
- INDEX
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Basis of Progressive Evolution by G. Ledyard Stebbins in PDF and/or ePUB format, as well as other popular books in Scienze biologiche & Scienza generale. We have over one million books available in our catalogue for you to explore.