Drawn from a 2005 international symposium, these essays explore current tyrannosaurid current research and discoveries regarding Tyrannosaurus rex.
The opening of an exhibit focused on "Jane," a beautifully preserved tyrannosaur collected by the Burpee Museum of Natural History, was the occasion for an international symposium on tyrannosaur paleobiology. This volume, drawn from the symposium, includes studies of the tyrannosaurids
Chingkankousaurus fragilis and "Sir William" and the generic status of
Nanotyrannus; theropod teeth, pedal proportions, brain size, and craniocervical function; soft tissue reconstruction, including that of "Jane"; paleopathology and tyrannosaurid claws; dating the "Jane" site; and tyrannosaur feeding and hunting strategies.
Tyrannosaurid Paleobiology highlights the far ranging and vital state of current tyrannosaurid dinosaur research and discovery.
"Despite being discovered over 100 years ago, Tyrannosaurus rex and its kin still inspire researchers to ask fundamental questions about what the best known dinosaur was like as a living, breathing animal.
Tyrannosaurid Paleobiology present a series of wide-ranging and innovative studies that cover diverse topics such as how tyrannosaurs attacked and dismembered prey, the shapes and sizes of feet and brains, and what sorts of injuries individuals sustained and lived with. There are also examinations of the diversity of tyrannosaurs, determinations of exactly when different kinds lived and died, and what goes into making a museum exhibit featuring tyrannosaurs. This volume clearly shows that there is much more to the study of dinosaurs than just digging up and cataloguing old bones." —Donald M. Henderson, Royal Tyrrell Museum of Palaeontology

eBook - ePub
Tyrannosaurid Paleobiology
- 312 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
Physical SciencesSubtopic
Biology1
Systematics and Descriptions
1
Phylogenetic Revision of Chingkankousaurus fragilis, a Forgotten Tyrannosauroid from the Late Cretaceous of China
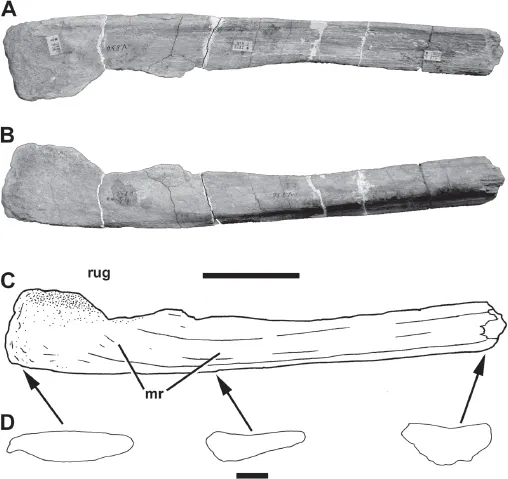
1.1. Photographs and line drawings of the holotype of Chingkankousaurus fragilis Young, 1958 (IVPP V 836, right scapula). A) Photograph in lateral view (dorsal to top). B) Photograph in medial view (dorsal to bottom). C) Line drawing in medial view (dorsal to bottom). D) Cross sections from the three indicated areas (lateral to top). Abbreviations: mr, medial ridge; rug, ruosities on posterior expansion of blade. Top scale bar equals 10 cm; bottom scale bar (for cross sections) equals 2 cm.
Introduction
Recent discoveries, especially the feathered theropods of the Jehol Biota, have placed China at the forefront of contemporary dinosaur research (e.g., Chen et al. 1998; Xu et al. 2003; Norell and Xu 2005; Xu and Norell 2006). However, vertebrate paleontology has a long history in China, and the country’s rich dinosaur fossil record has been explored for over a century. Much of the pioneering work on China’s dinosaurs was led by C. C. Young (Yang Zhongjian), the “father of Chinese vertebrate paleontology.” For over 40 years, from the early 1930s until his death in 1979, Young spearheaded expeditions across China and discovered many of the country’s most recognizable dinosaurs, such as the colossal sauropod Mamenchisaurus and the prosauropods Lufengosaurus and Yunnanosaurus (Dong 1992).
In 1958, Young described a single fragmentary bone from the Late Cretaceous (Campanian-?Maastrichtian; see Weishampel et al. 2004; Zhao et al. 2008) Wangshi Series of Shandong Province as a new genus and species of giant theropod, Chingkankousaurus fragilis. This specimen, the posterior region of a large right scapula (IVPP V 836), has long been ignored because of its fragmentary condition. However, those authors who have considered this specimen have often disagreed about its phylogenetic affinities. Young himself (1958) noted similarities with Allosaurus, and much later Dong (1992) formally assigned the specimen to Allosauridae. Steel (1970) and Dong (1979) placed the specimen within Megalosauridae, a wastebasket assemblage of large theropods that are now regarded as basal tetanurans (Benson 2010; Benson et al. 2010). Finally, Molnar et al. (1990:199) referred IVPP V 836 to Tyrannosauridae “on the basis of its very slender scapular blade.” This referral was taken one step further by Holtz (2004), who synonymized Chingkankousaurus with the common Asian Late Cretaceous tyrannosaurid Tarbosaurus. Unfortunately, most of these referrals have been based on vague criteria and were often simply asserted instead of supported by explicit discussion of characters and measurements. This was often unavoidable at the time, but an influx of new theropod discoveries from Asia and elsewhere over the past two decades now allows a firm basis for comparison.
In this chapter, we reassess IVPP V 836 based on firsthand examination of the specimen, compare it with the scapulae of other theropods, and use this information to comment on the taxonomy and phylogenetic placement of Chingkankousaurus fragilis. Although a systematic revision of a fragmentary specimen may seem trivial, it is important to establish the phylogenetic affinities of IVPP V 836 because this specimen has been referred to many disparate theropod groups and comes from an area (Shandong) where the theropod fauna has been more poorly sampled than in many other regions in China. If it truly does represent an allosauroid or megalosaurid, then this specimen would be among the last surviving members of these groups, would greatly expand their stratigraphic ranges in Asia, and would indicate that more basal theropods persisted alongside tyrannosaurids in the large predator niche of Late Cretaceous Asia (contrary to Brusatte et al. 2009b). However, if IVPP V 836 represents a tyrannosaurid or a closely related form, it is further evidence that that these enormous theropods were the sole large predators during the waning years of the Cretaceous in Laurasia.
Institutional Abbreviations AMNH, American Museum of Natural History, New York; HMB, Humboldt Museum für Naturkunde, Berlin; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing; JME, Jura Museum, Eichstatt, Germany; LH, Long Hao Institute of Geology and Paleontology, Hohhot, China; MCNA, Museo de Ciencias Naturales y Antropológicas (J. C. Moyano) de Mendoza, Mendoza, Argentina; MPC, Mongolian Paleontological Center, Ulaanbaatar; UMNH, Utah Museum of Natural History, Salt Lake City.
Phylogenetic Definitions and Phylogenetic Framework
In this chapter we use the phylogenetic definitions of Sereno et al. (2005) for Tyrannosauroidea and Tyrannosauridae. Tyrannosauroidea is defined as the most inclusive clade containing Tyrannosaurus rex but not Ornithomimus edmontonicus, Troodon formosus, or Velociraptor mongoliensis. The more derived Tyrannosauridae is defined as the least inclusive clade containing T. rex, Gorgosaurus libratus, and Albertosaurus sarcophagus. In our discussion of tyrannosauroid phylogeny, we follow the phylogenetic analysis and cladogram presented by Brusatte et al. (2010). This cladogram is depicted in Figure 1.5, and major clades are denoted.
Identification
Although fragmentary, IVPP V 836 (Fig. 1.1) can be identified as a partial right scapula owing to its shape and features of its morphology. This bone was originally described as a scapula by Young (1958), an identification that has been followed by subsequent authors (e.g., Molnar et al. 1990). However, Chure (2000) questioned this identification, noting that the symmetrical cross section figured by Young (1958) is unusual for a scapula. Although Young (1958) describes the cross section as symmetrical, in fact the medial surface is convex, and the lateral surface is flat to slightly concave, as is usual for theropod scapulae (Fig. 1.2). This results in a triangular cross section at mid-shaft and a semi-ovoid cross section anteriorly at the broken edge (Fig. 1.1D). The medial convexity is due to a pronounced ridge, described below, which is a normal feature for tyrannosaurid (e.g., Brochu 2003:fig. 80) and other theropod scapulae (Fig. 1.2A–B). Other features of the bone, such as the slightly concave lateral surface and weakly rugose distal end, are also present in theropod scapulae (Fig. 1.2C–D).
Other possible identifications for the bone, including the possibility that it is part of a dorsal rib or a gastral element, are untenable. The specimen is straight along its entire length, whereas theropod dorsal ribs are strongly curved, and only very small fragments would appear straight if observed in isolation (e.g., Madsen 1976:pl. 40; Brochu 2003:fig. 64). Additionally, the dorsal ribs of large theropods often bear a thick ridge on their anterior surface, which is paralleled by a depressed groove (e.g., Daspletosaurus: AMNH 5468). The posterior surface is often corrugated, with a deep groove corresponding to the ridge on the lateral surface. This morphology is not present in IVPP V 836, which has a single ridge on one surface and a flat to slightly concave opposing surface. Although the distal ends of anterior dorsal ribs are sometimes expanded to articulate with the sternum, these expansions are usually slight and rarely, if ever, more than twice mid-shaft depth, as is the case in IVPP V 836 (e.g., Lambe 1917:figs. 6, 7; Brochu 2003:fig. 64).
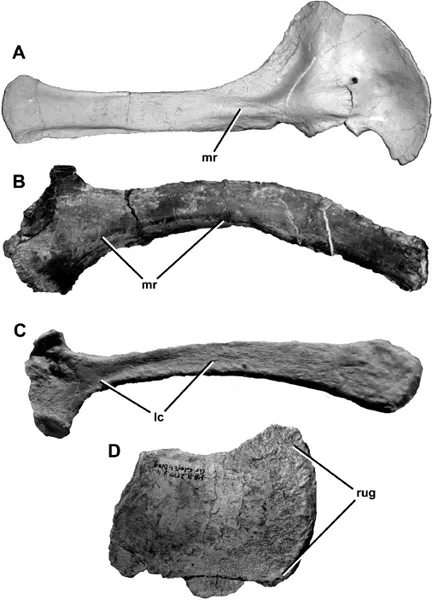
1.2. Comparative figure showing morphological features common to theropod scapulae, each of which is present on IVPP V 836, supporting its identification as a scapula. A) Aerosteon riocoloradensis (MCNA-PV-3137), left scapula in medial view. B) Falcarius utahensis (UMNH 12279), right scapula in medial view. C) Juravenator starki (JME Sch 200), left scapula in lateral view. D) Liliensternus liliensterni (HMB MB.R.2175), distal end of scapula in lateral or medial view. Abbreviations: lc, lateral concavity; mr, medial ridge; rug, rugosities on posterior expansion of blade.
Similarly, gastral elements of the largest theropods, such as Tyrannosaurus, are smaller than IVPP V 836, and their detailed morphology differs (e.g., Brochu 2003:fig. 70). In particular, although the medial ends of the medial gastral elements may be expanded relative to the mid-shaft, these expansions are usually irregular in shape (not spatulate as in IVPP V 836), extremely rugose, and often fused to the opposing medial gastral element. Additionally, IVPP V 836 is extremely large for a theropod gastral element.
Description
IVPP V 836 is the posterior end of a right scapula, measuring 520 mm long anteroposteriorly as preserved (Fig. 1.1). It is 47 mm deep dorsoventrally at its broken proximal end, and it maintains a relatively constant depth for most of the length of the shaft. However, it thins slightly to 43 mm in depth before expanding distally into a spatulate shape. As preserved, this expansion is 83 mm deep, but both its dorsal and ventral margins are eroded. When the preserved dorsal and ventral margins of the more proximal shaft are extended distally, filling in some of the missing regions, it appears as if the distal expansion was at least 94 mm deep. It is likely, however, that it was even deeper in life, as both the dorsal and ventral edges of the expansion are still quite thick, whereas they usua...
Table of contents
- Cover
- Half title
- Title
- Copyright
- Contents
- Contributors
- Introduction
- Part 1 Systematics and Descriptions
- Part 2 Functional Morphology and Reconstruction
- Part 3 Paleopathology, Paleoecology, and Taphonomy
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Tyrannosaurid Paleobiology by J. Michael Parrish, Ralph E. Molnar, Philip J. Currie, Eva B. Koppelhus, J. Michael Parrish,Ralph E. Molnar,Philip J. Currie,Eva B. Koppelhus, RALPH MOLNAR, PHILIP J CURRIE, EVA B KOPPELHUS in PDF and/or ePUB format, as well as other popular books in Physical Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.