An in-depth look at the origin and evolutionary radiation of the synapsids.
About 320 million years ago a group of reptiles known as the synapsids emerged and forever changed Earth's ecological landscapes. This book discusses the origin and radiation of the synapsids from their sail-backed pelycosaur ancestor to their diverse descendants, the therapsids or mammal-like reptiles, that eventually gave rise to mammals. It further showcases the remarkable evolutionary history of the synapsids in the Karoo Basin of South Africa and the environments that existed at the time. By highlighting studies of synapsid bone microstructure, it offers a unique perspective of how such studies are utilized to reconstruct various aspects of biology, such as growth dynamics, biomechanical function, and the attainment of sexual and skeletal maturity. A series of chapters outline the radiation and phylogenetic relationships of major synapsid lineages and provide direct insight into how bone histological analyses have led to an appreciation of these enigmatic animals as once-living creatures. The penultimate chapter examines the early radiation of mammals from their nonmammalian cynodont ancestors, and the book concludes by engaging the intriguing question of when and where endothermy evolved among the therapsids.
"Ever since Nick Hotton's book from the 1980s we have needed an update on the biology of therapsids, and it has been Anusuya Chinsamy-Turan and her students and associates who through their bone histological work have made the greatest progress in this field." —Martin Sander, Steinmann Institute, University of Bonn
"
Forerunners of Mammals is full of meticulous detail . . . [I]t also contains a number of excellently rendered illustrations of some of the animals covered in the book, and the final chapter is a discussion of the evolution of endothermy that anyone with a background in biology might find of interest. . . . Recommended." —
Choice
"
Forerunners of Mammals will take interested readers beyond the classic jaw-to-ear appreciation of therapsids, towards a deeper appreciation of the ancestry of mammals." —
Journal of Mammalian Evolution
"This volume represents a state-of-the-art contribution to our understanding of the paleobiology of how mammals arose, and what factors contributed to their evolutionary radiation and eventual success. It is highly recommended for anyone interested in these topics, and will be accessible to readers with minimal background in bone histology and synapsid paleontology." —
Quarterly Review of Biology

- 352 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
Physical SciencesSubtopic
Biology1
The Origin and Radiation of Therapsids
Tom S. Kemp
Introduction
The earliest fossils of amniotes—the clade that now consists of the reptiles birds and mammals—occur in 320 million-year-old rocks of the Late Carboniferous (sensu Laurin 2004; Laurin and Reisz 1995; Voigt and Ganzelewski 2010). They are characterized by several modifications that indicate an increased independence of freestanding bodies of water. For example, the aquatic sensory system of lateral lines is no longer present, the skull and jaws are strengthened for the biting action that terrestrial animals tend to rely on, and the limbs and girdles are robust. Whether they possessed the single, most characteristic of all amniote characters, an amniotic egg capable of developing on dry land, is unknown but seems probable (Packard and Seymour 1997). Surprisingly perhaps, the divergence between the two major living amniote sister groups was in existence from the very beginning of the fossil record of amniotes (Laurin and Reisz 1995). The Sauropsida consists of the modern reptiles and birds, plus the great range of dinosaurs, pterosaurs, and aquatic reptiles that dominated the land, sea, and air of the Mesozoic. The second group, the Synapsida, consists nowadays only of the mammals with their high-energy, active lifestyle and extraordinary ability to adapt successfully to so many terrestrial, subterranean, arboreal, aquatic, and aerial habitats in such a wide range of environmental conditions.
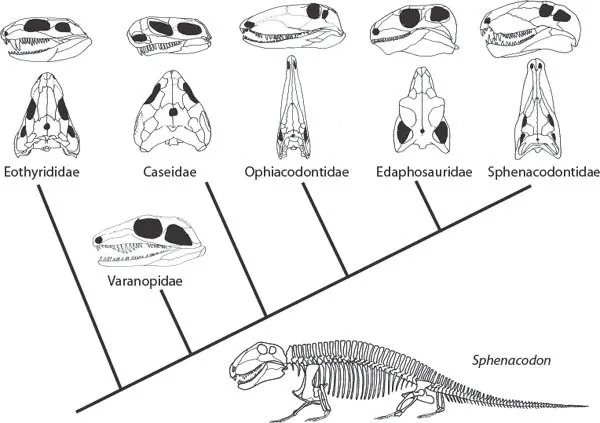
1.1. Phylogeny of the pelycosaurian-grade synapsids (reproduced by permission of Oxford University Press from “Interrelationships of the Synapsida” by Kemp [1988]), with a reconstruction of the skeleton of the sphenacodontid pelycosaur Sphenacodon (redrawn from Romer and Price [1940]).
The connection between the mammals and their remote Carboniferous relatives is one of the most remarkable parts of the fossil record. These animals, referred to as nonmammalian synapsids, or more affectionately if a trifle misleadingly as “mammal-like reptiles,” are the stem-group mammals, and they exhibit a range of different combinations of basal amniote and mammalian characters. Most of these nonmammalian synapsids are included in Therapsida, the monophyletic taxon that originated within a primitive, paraphyletic group called “pelycosaurs,” at least by the Middle Permian when the first undisputed therapsids are found. The therapsids are of fundamental importance in the history of terrestrial life on Earth for three reasons: first, because they dominated the terrestrial vertebrate scene from the Middle through the Late Permian and remained one of the most significant groups of the Triassic; second, because they established for the first time a terrestrial ecosystem based on very large numbers of fully terrestrial herbivorous tetrapods as the primary consumers, plus related carnivores as the major secondary consumers, a pattern to be repeated later by the dinosaurs of the Mesozoic and the mammals of the Tertiary; and third, because it was the therapsids that commenced the evolution of an elevated energy budget, and therefore a lifestyle in which relative independence of environmental fluctuations in temperature allowed continuous high levels of activity. Progressive development of this revolutionary biological strategy can be followed all the way to its expression in the fully endothermic mammals (Kemp 2006b, 2007b).
The most recent overall review of the evolution of the synapsids is that of Kemp (2005). Reisz (1986) reviewed the pelycosaurs to genus level. Rubidge and Sidor (2001), and somewhat egregiously Ivakhnenko (2003), reviewed the major therapsid subtaxa.
The Origin and Early Radiation of the Synapsida: Grade “Pelycosauria”
Synapsida is a monophyletic taxon recognized by a number of characters, including most distinctively the eponymous synapsid temporal fenestra in the skull roof, lying behind the orbit, and primitively bounded by the postparietal, parietal, squamosal, quadratojugal and jugal bones of the skull. The earliest record of synapsids consists of footprints of early Late Carboniferous age found in Germany, which predates the oldest actual fossil synapsids by 5–10 Ma (Voigt and Ganzelewski 2010). The latter consists of the humerus and other fragments described as Protoclepsydrops by Carroll (1964), which are dated as Westphalian B of the Late Carboniferous of Nova Scotia, and the slightly younger Westphalian D ophiacodontid pelycosaur Archaeothyris from the same region, which is represented by a skull lacking the mandible, and by the vertebral column and parts of the limbs (Reisz 1972).
The Late Carboniferous and Early Permian witnessed a radiation of nontherapsid synapsids, usefully referred to as the paraphyletic group “Pelycosauria.” They retained many unmodified amniote features such as a more or less homodont dentition, short and heavily built sprawling limbs, and a long massive tail (Fig. 1.1).
The great majority are known from North America, although there are also representatives in Europe and, at the very end of their temporal range, rare representatives of the family Varanopidae in South Africa (Modesto et al. 2001; Botha-Brink and Modesto 2009). In their classic review, Romer and Price (1940) divided all pelycosaurs into three major subtaxa, the primitive, mostly long-snouted piscivorous Ophiacodontia, the herbivorous Edaphosauria, and the carnivorous Sphenacodontia. Reisz (1986) reduced these three taxa to family status, and recognized the more basal position of certain forms (Fig. 1.1), an arrangement largely supported by subsequent studies (e.g., Berman et al. 1995; Reisz, Godfrey, and Scott 2009; see chapter 4 of this book). The Early Permian Eothyris and Oedaleops are each known from a single, small skull and constitute a clade Eothyrididae that had retained the primitively broad form, large supratemporal bone in the temporal region, and carnivorous dentition (Reisz, Godfrey, and Scott 2009). The family Caseidae is related to the eothyridids and like them had a broad skull roof (Maddin, Sidor, and Reisz 2008). However, caseids were highly specialized for an herbivorous diet, exhibiting shortening of the skull and an enlarged temporal fenestra indicating powerful adductor musculature. The marginal dentition consists of closely packed leaf-like teeth and there is also a well-developed palatal dentition against which a tough tongue is presumed to have worked. Although low on the phylogenetic tree, caseids are actually the last major group to appear in the fossil record and were also one of the two last representatives of pelycosaurian-grade animals to survive, being still present in the Middle Permian of Russia.
All other pelycosaurs are included in the monophyletic clade Eupelycosauria, distinguished by narrowing of the skull and a reduction of the supratemporal bone. Varanopids were relatively small, carnivorous forms, and there are three derived eupelycosaurian groups, each specialized for a different basic mode of life. The Ophiacodontidae (which include Archaeothyris) are long-snouted forms with a marginal row of small, sharp teeth suitable for piscivory. The Edaphosauridae have a number of features for herbivory that are convergent on the caseids, such as short jaws, large temporal fenestra, and a battery of palatal teeth that in this case was opposed by a similar battery on the medial side of each mandible. The edaphosaurid postcranial skeleton is distinguished by enormously elongated neural spines bearing short crosspieces or tubercles (see Plate A). The presumed soft tissue sail that these supported could hardly fail to have had the potential to enhance ectothermic temperature regulation of these animals (Bennett 1996), although others have argued that they were primarily related to sexual selection (Tomkins et al. 2010). The final taxon is the carnivorous Sphenacodontidae, most famously Dimetrodon (see Plate A; Fig. 4.5C), with its caniniform teeth and a body up to three meters in length. It too possesses elongated neural spines, in this case lacking the crosspieces and no doubt evolved independently of those of edaphosaurids. Sphenacodon possesses much less elongated neural spines, and Secodontosaurus is a curiously long-snouted, piscivorous form, although Reisz, Berman, and Scott (1992) suggested that such a sail-bearing tetrapod was more likely a specialized insectivore or carnivore taking its food from crevices.
As far as is presently known, pelycosaurs were restricted during the Late Carboniferous and Early Permian to the equatorial latitudes of central Pangaea, a warm, permanently humid biome, which suggests that during this period they were unable to survive excessively arid or cool conditions (Kemp 2006b). The very few caseids and varanopids that lingered into the Middle Permian occur in higher latitudes and may perhaps have evolved seasonal torpor strategies to survive the less equable conditions they met. There was also, paradoxically, a far higher proportion of carnivorous to herbivorous pelycosaurs than expected in a terrestrial tetrapod community, even among ectotherms with low metabolic rates. This can be accounted for by the hypothesis that the ecosystem was still dependent to a considerable extent on freshwater aquatic productivity (Milner 1993; Berman, Sumida, and Lombard 1997; Kemp 2006b; Olson 1975, 1986). The presence of abundant fish-eating ophiacodontids and perhaps Secodontosaurus as intermediates in the energy flow from freshwater to dry land is certainly consistent with this view.
The Origin and Interrelationships of Therapsids
The origin of the therapsids by the Middle Permian was a revolution in the history of terrestrial life, because they show clear morphological evidence of higher activity levels and, therefore, presumably higher energy budgets (reviewed in Kemp [2005]; see chapter 11 of this book). The evolutionary changes in the skull mainly concern the mechanics of feeding. Enlargement of the temporal fenestra and a correlated enlargement of the coronoid region of the mandible indicate enlarged, more strongly attached adductor mandibuli musculature. This is associated with a forward shift of the jaw articulation, which shortens the jaw and so increases the bite force, and a general strengthening of the posterior part of the skull. Clearly this stronger bite was associated with the considerably enlarged canine teeth and well-developed, often interdigitating incisors. There was an equally significant transition in the postcranial skeleton, from the stout-limbs and sprawling gait of the pelycosaurs to longer and more gracile limbs and increased mobility at the shoulder and hip joints of therapsids (see Fig. 1.5D). The new gait appears to have endowed therapsids with a great deal more agility as well as speed (Gebauer 2007; Kemp 1978), and by raising the body clear of the ground, it increased the potential ventilation capacity of the lungs (Carrier 1987). The bone histology of therapsids also offers evidence for a more active, higher energy mode of life, as discussed in later chapters of this book. Much of the therapsid bone is fibrolamellar, which indicates a relatively high rate of bone deposition and growth (Botha and Chinsamy 2004, 2005; Ray, Botha, and Chinsamy 2004; Ray and Chinsamy 2004), although how exactly this relates to the evolution of temperature physiology remains debatable. For example, there is a good deal of variation even among related genera in the extent to which growth was continuous or interrupted during the unfavorable season, when food would have been in short supply.
It is surely no coincidence that therapsids make their first appearance in temperate paleolatitudes where seasonality was pronounced, suggesting that there had already been a significant degree of evolution of the temperature and osmotic regulatory strategies that were to culminate in those of the mammals (Kemp 2006a, 2006b). Apart from one or two briefly lingering forms, the pelycosaurs disappeared at this time, leaving the therapsids as the dominant terrestrial tetrapods of the Late Permian. There was also a significant shift toward a higher proportion of herbivores to carnivores (Nicolas and Rubidge 2009; Sidor and Smith 2007; see chapter 2 of this book), indicating the development of a community structure analogous to that of today’s mammals, with virtually complete dependence on land plants as the primary producers and herbivorous tetrapods as the primary consumers.
The phylogenetic relationships of Therapsida are indicated by several features shared only with the sphenacodontid pelycosaurs, of which the most memorable is a reflected lamina of the angular of the lower jaw. This is little more than a notch in the ventral edge of the back of the jaw in sphenacodontids, but has become a large, thin lateral sheet in therapsids, whose function continues to be unclear. One view is that it represents the attachment of ventral jaw musculature, the other that it was involved in airborne sound reception, as an incipient stage in the origin of mammalian hearing (Allin 1975). Other similarities are the enlarged caniniform teeth and the powerful build of the occiput.
Several subtaxa of therapsids appeared approximately simultaneously around 265–270 Ma in Guadalupian aged rocks of South Africa, Russia, and China (Abdala, Rubidge, and van den Heever 2008; Fröbisch 2009). Together they constitute a very strongly supported monophyletic taxon, characterized by many features of the skull and dentition related to feeding, such as the enlarged canines and enlarged temporal fenestra, and a major reorganization of the locomotory system involving longer, more gracile limbs and a greater range of movements at their joints. Little is yet known of more basal therapsids. Tetraceratops was described long ago on the basis of a single poorly preserved skull lacking lower jaws and dating from the Early Permian of Texas; Romer and Price (1940) interpreted it as possibly an eothyridid pelycosaur. However, when Laurin and Reisz (1996) restudied the specimen, they concluded that it possessed a number of therapsid characters such as the form of the temporal fenestra and the manner of attachment of the braincase. Others, however, remain skeptical that the specimen is adequate for such an interpretation (Liu, Rubidge, and Li 2009; Rubidge and Sidor 2001), and resolution of this potentially very important issue must await further material of the species. Apart from the p...
Table of contents
- Cover
- Half title
- Title
- Copyright
- Dedication
- Contents
- Preface
- Acknowledgments
- List of Contributors
- 1 The Origin and Radiation of Therapsids
- 2 Therapsid Biodiversity Patterns and Paleoenvironments of the Karoo Basin, South Africa
- 3 The Microstructure of Bones and Teeth of Nonmammalian Therapsids
- 4 The Paleobiology and Bone Microstructure of Pelycosaurian-Grade Synapsids
- 5 Dicynodont Growth Dynamics and Lifestyle Adaptations
- 6 Biological Inferences of the Cranial Microstructure of the Dicynodonts Oudenodon and Lystrosaurus
- 7 Bone and Dental Histology of Late Triassic Dicynodonts from North America
- 8 Bone Histology of Some Therocephalians and Gorgonopsians, and Evidence of Bone Degradation by Fungi
- 9 The Radiation and Osteohistology of Nonmammaliaform Cynodonts
- 10 The Radiation, Bone Histology, and Biology of Early Mammals
- 11 The Evolution of Mammalian Endothermy
- References
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Forerunners of Mammals by Anusuya Chinsamy-Turan in PDF and/or ePUB format, as well as other popular books in Physical Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.