This volume on iron-sulfur proteins includes chapters that discuss how microbes, plants, and animals synthesize these complex prosthetic groups, and why it is important to understand the chemistry and biogenesis of iron sulfur proteins. In addition to their vital importance in mitochondrial respiration, numerous iron sulfur proteins are important in maintenance of DNA integrity. Multiple rare human diseases with different clinical presentations are caused by mutations of genes in the iron sulfur cluster biogenesis pathway. Understanding iron sulfur proteins is important for understanding a rapidly expanding group of metabolic pathways important in all kingdoms of life, and for understanding processes ranging from nitrogen fixation to human disease.
Trusted by 375,005 students
Access to over 1 million titles for a fair monthly price.
1A retrospective on the discovery of [Fe-S] cluster biosynthetic machineries in Azotobacter vinelandii
Patricia C. Dos Santos and Dennis R. Dean
1.1Introduction
Iron-sulfur clusters ([Fe-S]) clusters are inorganic prosthetic groups that participate in many essential life processes. Despite their relatively simple structures, the biological assembly of [Fe-S] clusters is complicated and the defining features of the process have only emerged over the past two decades. The biosynthetic process involves two principle interacting proteins, an l-cysteine desulfurase and a molecular assembly scaffold. In this chapter, a retrospective is provided that describes the initial discovery of the mechanism for the assembly of [Fe-S] clusters in the nitrogen-fixing organism Azotobacter vinelandii. Why and how the study of biological nitrogen fixation ultimately contributed the key insights into the general process of [Fe-S] cluster assembly is described.
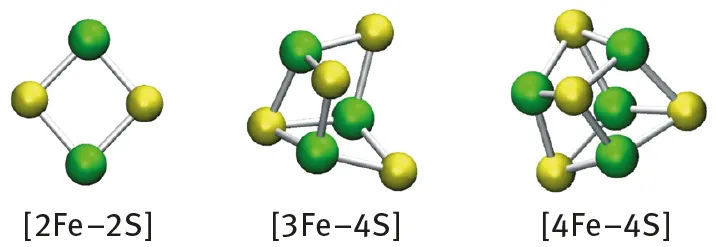
Beinert, Holm, and Munck, pioneers in the field of bioinorganic chemistry, have aptly described biological iron-sulfur clusters ([Fe-S] clusters) as nature’s modular, multipurpose structures [1]. What are [Fe-S] clusters, why do they exist in nature, and why are they so important to so many biological processes? [Fe-S] clusters were originally discovered as inorganic prosthetic groups composed exclusively of iron and sulfide contained in a class of redox-active proteins denominated as ferredoxins [2, 3]. Ferredoxins are electron/proton carrier proteins that contain [Fe-S] clusters, usually in the form of rhombic [2Fe-2S] or cubane [4Fe-4S] clusters (Fig. 1.1). [Fe-S] clusters are most often covalently attached to their cognate protein partners through cysteinyl thiolate ligation to their metal sites, although other ligation modes are also known to exist and not all of the metal sites are necessarily coordinated by a protein-donated ligand [4]. Ferredoxins represent a specialized class of a wide variety of proteins, now generically designated as [Fe-S] proteins, that contain one or more [Fe-S] clusters. It is the capacity of [Fe-S] clusters to exist in multiple oxidation states that endows [Fe-S] proteins with their ability to serve as electron/proton carriers. Indeed, the reversibility of redox properties of [Fe-S] proteins is an integral aspect of essential energy transducing processes in nitrogen fixation, photosynthesis, and respiration. The key involvement of [Fe-S] clusters in such life-sustaining processes is intimately linked to the wide range of redox potentials they can attain as a consequence of their respective polypeptide environments [5]. However, the ability of proteins to tune the redox potentials of their cognate [Fe-S] clusters is not the only feature of [Fe-S] clusters exploited by nature. The chemical and structural versatility of [Fe-S] clusters, uniquely achieved by combining the individual chemical properties of Fe and S, has enabled [Fe-S] proteins to fulfill other important biological roles including activation of substrates for chemical transformations, serving as environmental sensors, providing structural determinants within proteins, and functioning as agents of gene regulation [4, 6].
Fig. 1.1: Simple [Fe-S] cluster types found in nature.
Although [Fe-S] clusters play a central role in many biological processes, there is also a significant penalty associated with their use. Indeed, Fe2+ and S2−, necessary for both the assembly and disassembly of [Fe-S] clusters, are toxic to a wide variety of cellular processes in aerobic organisms. How then did [Fe-S] clusters become so pervasive in biology? The answer to this question is not known for certain but could be linked to the “iron-sulfur world” theory, which proposes that chemical reactions associated with formation of the organic building blocks necessary for the emergence of life on earth occurred on metal-sulfur surfaces in the highly reducing environment of prebiotic earth [7, 8]. Certain aspects of such chemistries might have been captured in primordial organisms in the form of [Fe-S] clusters, particularly when considering that facile “spontaneous” assembly of [Fe-S] clusters occurs under reducing conditions. This possibility finds some credence when it is considered that life likely originated in an anoxic environment in which free Fe2+ and free S2− did not pose the same risk as is associated with the oxygen-saturated environment that dominates life on earth today. For example, the potential for lethal formation of reactive oxygen species (ROS) through Fenton chemistry did not exist for emergent life forms. This luxury was eliminated, however, by the advent of oxygenic photosynthesis, which gradually converted the biosphere from a reducing environment to a primarily oxidizing environment. Given that fundamental life sustaining processes requiring [Fe-S] clusters were almost certainly well developed by the time photosynthesis emerged, the spontaneous assembly of [Fe-S] clusters from free Fe2+ and S2− could not possibly continue, even if that was the case in primordial life. What this means is that at some point during evolution and certainly upon transition of earth to an oxidizing atmosphere, living organisms needed to develop a way to construct [Fe-S] clusters such that free Fe2+ and free S2− were not required [9, 10]. In other words, these elements needed to be trafficked and combined in nontoxic forms or living organisms needed to evolve alternative strategies to replace the many functions supplied by [Fe-S] proteins.
Considering the importance of [Fe-S] clusters in sustaining essential life processes [11] as well as the striking structural simplicity of most [Fe-S] clusters, it might seem curious that so little was known about their biological assembly until relatively recently. However, as is discussed in this narrative, there are very good reasons why a fundamental understanding of the assembly of [Fe-S] clusters took so long to develop when compared with our understanding of the biosynthetic pathways for formation of many other organic cofactors. One reason is that the critical importance of [Fe-S] clusters to so many biological processes prevented fortuitous discovery of assembly factors in genetic screens because assembly factors were necessary for survival. Another reason is that the structural simplicity of [Fe-S] clusters, and the ability to form them spontaneously in situ from free Fe2+ and S2−, failed to inspire serious inquiry into potential mechanisms for their biological formation. In this chapter, we provide a retrospective about the work using the bacterium Azotobacter vinelandii, and studies on the specialized process of biological N2 fixation, that ultimately led to some of the key insights about the [Fe-S] cluster assembly process.
1.2An introduction to nitrogenase
Serendipity is defined as the accident of finding something good or useful while not specifically searching for it. Initial work on nitrogenase was not aimed at understanding how simple [Fe-S] clusters are formed in biological systems. Instead, the focus was directed at understanding the genetic determinants and chemical mechanism of biological N2 fixation, an essential contributor to the biogeochemical nitrogen cycle. Here we discuss some of the general features of the enzyme nitrogenase, the catalytic component of N2 fixation, and the initial approaches that were used to explore its function [12, 13]. It will be seen that, by both chance and design, these approaches inexorably led to the discovery of unifying features involved in the assembly of simple [Fe-S] clusters necessary to sustain cellular metabolism.
Simple rhombic [2Fe-2S] and cubane [4Fe-4S] clusters (Fig. 1.1), composed only of Fe and S, represent the dominant forms of [Fe-S] clusters in biological systems [4]. However, there are also other types of [Fe-S] clusters that have higher nuclearity, those that contain another metal in addition to Fe, and those that also contain organic constituents [13, 14]. A broad spectrum of [Fe-S] cluster types can be found in nitrogenase, the enzyme that catalyzes the nucleotide-dependent reduction of N2 to yield two molecules of ammonia (NH3) (Fig. 1.2). Nitrogenase is a two component enzyme that contains a canonical [4Fe-4S] cluster, involved in electron transfer, a novel [8Fe-7S] cluster (P-cluster) that also serves as an agent of electron transfer, and a [7Fe-9S-Mo-C-homocitrate] cluster (FeMo cofactor, or FeMoco), which contains molybdenum (Mo) and provides the site for N2 activation and reduction [1...
Table of contents
Cover
Title Page
Copyright
Preface
Tracey A. Rouault biography
Contents
List of contributing authors
1 A retrospective on the discovery of [Fe-S] cluster biosynthetic machineries in Azotobacter vinelandii
2 The ISC system and the different facets of Fe-S biology in bacteria
3 A stress-responsive Fe-S cluster biogenesis system in bacteria – the suf operon of Gammaproteobacteria
4 Sensing the cellular Fe-S cluster demand: a structural, functional, and phylogenetic overview of Escherichia coli IscR
5 Fe-S assembly in Gram-positive bacteria
6 Fe-S cluster assembly and regulation in yeast
7 The role of Fe-S clusters in regulation of yeast iron homeostasis
8 Biogenesis of Fe-S proteins in mammals
9 Delivery of iron-sulfur clusters to recipient proteins: the role of chaperone and cochaperone proteins
10 Iron-sulfur proteins and human diseases
11 Friedreich ataxia
12 Connecting the biosynthesis of the molybdenum cofactor, Fe-S clusters, and tRNA thiolation in humans
13 Iron-sulphur proteins and genome stability
14 Eukaryotic iron-sulfur protein biogenesis and its role in maintaining genomic integrity
15 DNA signaling by iron-sulfur cluster proteins
16 Iron-sulfur cluster assembly in plants
17 Origin and evolution of Fe-S proteins and enzymes
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Biochemistry, Biosynthesis and Human Diseases by Tracey Rouault in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Microbiology. We have over one million books available in our catalogue for you to explore.