Two of the greatest evolutionary events in the history of life on Earth occurred during Early Paleozoic time. The first was the Cambrian explosion of skeletonized marine animals about 540 million years ago. The second was the "Great Ordovician Biodiversification Event," which is the focus of this book. During the 46-million-year Ordovician Period (489–443 m.y.), a bewildering array of adaptive radiations of "Paleozoic- and Modern-type" biotas appeared in marine habitats, the first animals (arthropods) walked on land, and the first non-vascular bryophyte-like plants (based on their cryptospore record) colonized terrestrial areas with damp environments.
This book represents a compilation by a large team of Ordovician specialists from around the world, who have enthusiastically cooperated to produce this first globally orientated, internationally sponsored IGCP (International Geological Correlation Program) project on Ordovician biotas. The major part is an assembly of genus- and species-level diversity data for the many Ordovician fossil groups. The book also presents an evaluation of how each group diversified through Ordovician time, with assessments of patterns of change and rates of origination and extinction. As such, it will become the standard work and data source for biotic studies on the Ordovician Period.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
The historical scientist focuses on detailed particulars—one funny thing after another—because their coordination and comparison permits us, by consilience of induction, to explain the past with… confidence (if the evidence is good).
STEPHEN JAY GOULD, Wonderful Life
This introductory chapter presents the scope, aims, and organization of the volume; outlines previous work on Ordovician biodiversity topics; and gives an overview of the chapters in the volume, from those briefly appraising the Ordovician World to those more comprehensively surveying the diversification patterns of the main Ordovician taxonomic groups. The chapter ends with some closing remarks on the Ordovician Radiation and future directions.
Scope and Aims
Two of the greatest evolutionary events in the history of life on earth occurred during Early Paleozoic time. The first was the Cambrian explosion of skeletonized marine animals—what Wilson (1992: 188) called the “big bang of animal evolution”—about 540 million years (m.y.) ago. The second was the “Great Ordovician Biodiversification Event,” the focus of this book. During the 46-m.y. span of Ordovician time (489–443 m.y. ago), an extraordinarily varied range of evolutionary radiations of “Cambrian-, Paleozoic- and Modern-type” biotas appeared, the most diversified occurring in marine continental platform to open ocean habitats. Animals (arthropods) also first walked on land, and based on their cryptospore record, the first nonvascular bryophytelike plants colonized damp areas on land.
The Cambrian explosion of skeletonized animals is now comparatively well documented, though the timing of the initial event still needs to be reassessed (Bowring and Erwin 1998). Books have been published on a number of aspects of this explosion, for example, Glaessner’s The Dawn of Animal Life: A Biohistorical Study (1984), Lipps and Signor’s Origin and Early Evolution of the Metazoa (1992), and Zhuravlev and Riding’s The Ecology of the Cambrian Radiation (2000). Also well documented is the history of the extraordinary Mid Cambrian Burgess Shale faunas, about 505 m.y. ago. The most notable publications are Gould’s Wonderful Life (1989), Briggs, Erwin, and Collier’s The Fossils of the Burgess Shale (1994), and Conway Morris’s The Crucible of Creation (1998b).
In contrast, until very recently, there was no Ordovician biodiversity volume, or at least none that focused significantly on aspects of Ordovician biodiversity. However, nearly half the contributions in Crame and Owen’s Palaeobiogeography and Biodiversity Change: The Ordovician and Mesozoic-Cenozoic Radiations (2002) have addressed issues relating to Ordovician diversity change. The important role of plate tectonics during the Ordovician was singled out—examples, such as the fragmentation of the Gondwanan margin, the drift of Avalonia, and the development of an array of marginal to oceanic terranes (including island chains) that effectively partitioned ocean circulation patterns within the Iapetus Ocean—as prominent in engendering the patterns of diversity change.
Ordovician biodiversity has typically been studied in three different ways: (1) taxonomic diversity, which involves a focus on the taxonomic richness and turnover (originations and extinctions) within fossil groups; (2) ecologic diversity, which examines how organisms (or their communities) adapt to fill niche spaces in order to exploit available food resources more successfully; and (3) morphological diversity (usually termed disparity), which traces the patterns of morphological (design) change in various fossil groups.
This book is devoted primarily to documenting the taxonomic diversification of Ordovician biotas, in both global and regional contexts, and to firming up the timing of the most important diversification events. The volume has a broad coverage, but inevitably there are gaps. A few aspects of ecologic diversity have been treated, but the coverage is limited, and aspects of morphological diversity (disparity) are barely touched on. Moreover, only limited discussion of possible causes for the radiation events is included.
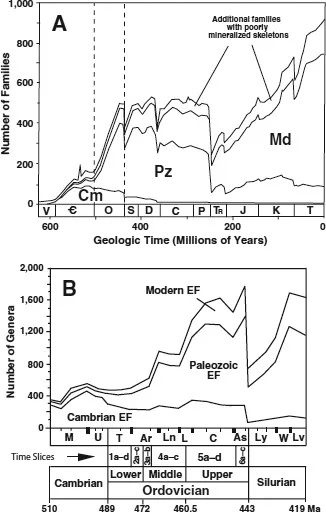
All these Ordovician diversification events form part of the “Great Ordovician Diversification Event,” also called the Ordovician Radiation (Droser et al. 1996). Although the most intensive part of the Ordovician Radiation was during the Mid to Late Ordovician epochs, an interval of 28 m.y. (until the second extinction pulse of the end Ordovician mass extinctions—Sheehan 2001b; chapter 9 in this volume), some taxonomic groups (e.g., trilobites, inarticulated brachiopods, graptolites, conodonts, and rostroconch mollusks) also diversified significantly during the Early Ordovician. Consequently, all these evolutionary events from the beginning to virtually the end of the Ordovician Period—through nearly 46 m.y. of earth history—should be treated as belonging to the Ordovician Radiation (figure 1.1).
FIGURE 1.1. (A) Phanerozoic taxonomic diversity of marine animal families (slightly modified from Sepkoski 1984: figure 1). The fields Cm, Pz, and Md represent the Cambrian, Paleozoic, and Modern “evolutionary faunas” (EFs), respectively. Note also the generalized field of additional, poorly preserved families at the top of the diagram. Sepkoski’s timescale follows Harland et al. (1982) and includes the following abbreviations: V = Vendian;
= Cambrian; O = Ordovician; S = Silurian; D = Devonian; C = Carboniferous; P = Permian; TR = Triassic; J = Jurassic; K = Cretaceous; T = Tertiary. (B) The Middle to Upper Cambrian, Ordovician, and Silurian (except Pridoli) taxonomic diversity of marine animal genera (modified from Sepkoski 1995: figure 1). The main field of Cambrian, Paleozoic, and Modern EFs is shown, as well as Sepkoski’s time units, including British series for his Ordovician and Silurian subdivisions. His abbreviations are as follows: M = Middle Cambrian; U = Upper Cambrian; T = Tremadocian; Ar = Arenig; Ln = Llanvirn; L = “Llandeilo”; C = Caradoc; As = Ashgill; Ly = Llandovery; W = Wenlock; Lv = Ludlow. The “Llandeilo” series has now been abandoned in favor of an enlarged Llanvirn, the name Llandeilian retained for its upper stage (figure 2.1), and the overlying Caradoc expanded downward to fill the gap (Fortey et al. 1995, 2000). Note also the added scale bar at the bottom of the diagram that comprises the tripartite global series subdivisions for the Ordovician System, radiometric dates in millions of years (Ma), and the time-slice subdivisions used in this volume.
No comparable, well-defined major extinction episode has been recognized at the beginning of the Ordovician Period. According to Sepkoski (1981b, 1995, 1997), after the dramatic Early Cambrian radiations there was a phase of apparent “stagnation” or quiescence that persisted through the Mid Cambrian to the Early Ordovician, with a comparatively lower overall diversity thoughout (figure 1.1B). Yet it remained an interval of high turnover—with high extinction rates limiting the overall buildup of diversity, including the well-defined successive pulses of mainly trilobite-based Mid to latest Cambrian “biomere” extinctions (Palmer 1979; Ludvigsen 1982; Zhuravlev 2000). The last of these more or less coincides with the base of the North American Ibexian (Ross et al. 1997), that is, three conodont zones below the base of the Ordovician (Cooper et al. 2001). A few extinction horizons have been identified within the Ordovician, but they all appear to be relatively minor pulses that have yet to be demonstrated as being truly global events. For example, Ji and Barnes (1993, 1996) documented an Early Ordovician (mid Tremadocian) conodont extinction event in Laurentia; Sepkoski (1992b, 1995, 1996) reported extinction horizons in the late Mid Ordovician (near the bottom and top of the Darriwilian, respectively); and Patzkowsky et al. (1997) identified a climatic event with associated Late Ordovician (mid Caradoc) brachiopod-dominated extinction in Laurentia. The last has wider, probably even global, importance, correlating with climatic and other changes—including the “late Keila” extinctions of chitinozoans, acritarchs, and ostracodes—in Baltoscandia (Kaljo et al. 1996; Ainsaar et al. 1999).
Biotas
This book was compiled by a large team of Ordovician specialists from around the world, under the aegis of the joint UNESCO and International Union of Geological Sciences (IUGS) geosciences initiative—the International Geological Correlation Programme (IGCP)—which supported IGCP project no. 410. The aim in establishing this first globally oriented, internationally sponsored IGCP project was to have the “Great Ordovician Biodiversification Event” comprehensively evaluated in a collective effort. When IGCP 410 became formally established in 1997, data collection and analysis of biotas were coordinated mainly on a regional basis, but in 1998 a separate study program was developed with a more constrained global approach on the major fossil groups. This became an evaluation of how each taxonomic group diversified through Ordovician time, with assessments of patterns of diversity change, and rates of origination and extinction, based on the assembly of genus- and species-level taxonomic diversity data of the various fossil groups. Leading specialists and their colleagues were invited to participate in the three-year work program and to attend a major meeting organized by Mary Droser in June 2001 at the University of California in Riverside to discuss the results of the compilations. The book derives in part from these contributions, with a number of additional studies from other specialists added to the project after the Riverside meeting to widen the coverage to include nearly all fossil groups.
The leader of each fossil group chose his or her own team of co-workers to tackle the compilation of data and assembly of the manuscript for the particular chapter. The groups varied in size from large (with a coordinating author and many coauthors) to small, single-author presentations. We encouraged each team to establish its diversity surveys so that they highlighted patterns of diversity change, originations, and extinctions, where possible down to species level at least for the pelagic groups, and to genus, and where possible to species level for benthic groups. Most authors assembled their primary taxonomic data using their own databases (spreadsheets or census lists).
The pelagic groups (e.g., graptolites, chitinozoans, radiolarians) and some benthic groups (e.g., bryozoans, sponges, stromatoporoids, echinoderms) are presented as species-level, or combined species- and genus-level, diversity surveys. Larger benthic groups (e.g., trilobites, brachiopods, gastropods, bivalves, nautiloids) have been surveyed mainly at the genus level at this stage. Although we attempted to provide the widest possible coverage of Ordovician biotas, it was inevitable that some unevenness in the levels of documentation and analysis would occur from chapter to chapter. For a variety of reasons, a number of groups are presented with rather incomplete global diversity analyses. This may be because (1) only part of the group’s Ordovician record has been treated; (2) reliable data of well-preserved and diagnostic material are available only from one or two regions in the world, so the survey does not have a global focus; (3) genus- and species-level assignments of a group are unreliable or at least need further revision before a worldwide analysis can be attempted; or (4) present-day expertise on a particular group is lacking. Consequently, for a few groups, only preliminary statements could be included, and in some cases these assessments remain predominantly regionally based.
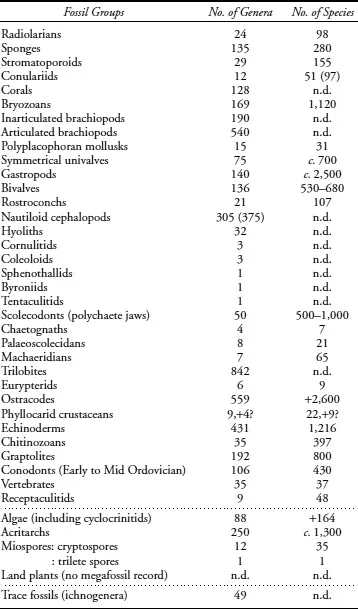
Although we have attempted to include most Ordovician groups in this genus- and species-level coverage (table 1.1), there are some gaps in the documentation—for instance, the Mid to Late Ordovician conodont diversity record and a few small groups of comparatively limited Ordovician occurrence, for example, foraminiferans (Lipps 1992a, 1992b), hydroids (Stanley 1986; Foster et al. 1999), and hyolithelminths. In addition, a few groups of microorganisms—such as two unicellular planktic green algal prasinophyte groups (leiospheres, tasmanitids) and the prolific, organic-walled, benthic cyanobacterium Gloeocapsomorpha prisca, a microorganism of considerable economic interest because it forms significant matlike accumulations of late Mid to Late Ordovician “kukersite” oil shales in the intracratonic basins of Estonia (Körts 1992), North America (Jacobson et al. 1988), and Australia (Foster et al. 1990)—have not been included in the survey.
An estimate of total numbers of Ordovician genera and species for each fossil group treated in the book (table 1.1) is provided by authors for general guidance only. It is not intended as a comprehensive listing of totals for all Ordovician groups. The list comprises about 4,605 genera (excluding the trace fossils). The total number of animal genera (less the “plants”—algae, acritarchs, miospores) is 4,254 genera. Earlier, Sepkoski (1995) employed a database of 4,367 animal genera (i.e., 12 percent of all known animal genera in the Phanerozoic) to prepare his outline of Ordovician diversity history. Only the few small groups mentioned earlier, the late Mid to Late Ordovician conodont record and about 70 genera of nautiloids from China and Russia, have been excluded from our present survey.
With recent publication of Sepkoski’s (2002) comprehensive listing of all known fossil marine animal genera, it is now possible to make more-up-to-date comparisons between the two lists of Ordovician animal generic diversity records (table 1.2). However, it is difficult to compare the two lists in detail because they represent completely different types of fossil data assemblies. The present study was not intended to be comprehensive but rather to involve as many available Ordovician specialists as possible in a stocktaking of their fossil group. In some cases these workers have adopted revised taxonomic categories from those listed in Sepkoski’s Compendium of Fossil Marine Animal Genera (2002). The Compendium represented a mammoth compilation of Phanerozoic animal genera, assembled from the data coverage of animal groups in Moore et al., Treatise on Invertebrate Paleontology (1953–1992), and many hundreds of other literature sources published up to 1998. Nevertheless, there is a remarkable degree of similarity between the two lists. The comprehensive Sepkoski (2002) compilation also provides a means to appreciate the full range of Ordovician taxonomic groups, including those biodiversity records that have not been documented here.
TABLE 1.1. A Preliminary Listing of Genus- and Species-Level Totals for the Ordovician Fossil Groups
Source: Compiled from data supplied by individual authors.
Note: This compilaton of animals, plants, and trace fossils is provided for general guidance only because some genus-level and especially the species-level taxonomy of fossil groups remains in a state of flux. For many groups a large backlog of taxonomic revision work still needs to be done.
Bivalve and scolecodont species data are shown as ranges, namely, between 530 and 680 and between 500 and 1,000 species, respectively. For entries of nautiloid genera and conulariid species, two numbers are included (one in parentheses): the lower number is the actual count (a mainly regional summation), and the higher number in parentheses is the global estimate. For the phyllocarids, the valid genera and species numbers are shown, plus the numbers of doubtful taxa (with a question mark). For trace fossils, only numbers of form genera (ichnogenera) could be supplied.
Abbreviations: c. = about; n.d. = data not determined (or not supplied).
TABLE 1.2. Comparative Generic Lists of Ordovician Animal Diversity Data
Source: Compiled from data supplied by individual authors (this study) and from Sepkoski (2002).
a Stromatoporoids are here listed (in this study and in Sepkoski’s list) as separate from other sponges.
b The problematic receptaculitids is the only group omitted from Sepkoski’s list. They are included in this study as a possible metazoan group.
c The bellerophontids have been included with the symmetrical univalves of this study, whereas they are included in Sepkoski’s list with the gastropods.
d The genus (Sphenothallus) is included in Sepkoski’s list as a conulariid.
e The genus (Byronia) is included in Sepkoski’s lists as a hyolithelminth.
Only a comparatively few groups have good-quality assignments of species data (table 1.1). These include mainly pelagic groups such as the graptolites, conodonts, chitinozoans, and radiolarians, but correct species assignments have also been established for a few benthic groups (e.g., stromatoporoids, conulariids, bryozoans, rostroconchs, eurypterids, echinoderms, and receptaculitids).
Wilson (1992) has an estimate of 1.4 million organisms (plants, animals, and microorganisms) currently known to be living on earth, though millions of insects, microbes, and other organisms remain undescribed across habitats from rain forests to the ocean deeps (Wilson 1992; Thorne-Miller 1999). The problems of estimating fossil species numbers is a more daunting task. Paul (1998) reviewed the available approaches, namely, to establish what proportion of living animal species is likely to be preserved in modern settings, as well as to establish the relationships between soft-bodied and skeletonized organisms in fossil Lagerstättern such as the Middle Cambrian Burgess Shale (Conway Morris 1986). These two approaches produce rather similar results, suggesting to Paul that only about 10 percent of all Phanerozoic species are likely to have been preserved. In assessing the Ordovician global record, we currently have only reasonably complete numbers for the genera—some 4,600 known Ordovician genera. From the generic and specific data in the 26 listed fossil groups (table 1.1), there are on average about five species for each genus. Multiplying this value by the generic total gives the very approximate total of 23,000 Ordovician skeletonized species. If this estimate represents on...
Table of contents
Cover
Half title
Series Page
Title
Copyright
Dedication
Contents
Acknowledgments
1. Introduction
Part I: Scaling of Ordovician Time and Measures for Assessing Biodiversity Change
Part II: Conspectus of the Ordovician World
Part III: Taxonomic Groups
Part IV: Aspects of the Ordovician Radiation
List of Figures and Tables
References
List of Contributors
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Great Ordovician Biodiversification Event by Barry D. Webby,Florentin Paris,Mary L. Droser,Ian G. Percival, Barry Webby, Florentin Paris, Mary Droser, Ian Percival in PDF and/or ePUB format, as well as other popular books in Scienze fisiche & Evoluzione. We have over 1.5 million books available in our catalogue for you to explore.