The southwestern Australian flora is unique in the world, not only for its biodiversity and endemism, but also for its functional biodiversity. It also contains the world's most nutrient-impoverished soils, has a prolonged-summer period and the vegetation is extremely fire-prone. These conditions have engendered an array of survival adaptations that have evolved in these harsh conditions across a diverse range of species. It is well recognised that the southwest flora has the toughest and most spiny vegetation of the world, the greatest number of species that store their seeds in woody fruits, and the most specialised means of obtaining limited soil nutrients and water.
This book focuses on the survival mechanisms, adaptations and ecology of the unique Southwest Australian flora (restricted here to flowering plants). The book begins with an examination of how the flora has evolved into the present forms. It describes further in detail the adaptive responses of the flora to the main environmental pressures influencing survival - fire, summer drought, nutrient-impoverished soils, pollination and seed dispersal agents. Specialised responses to obtain essential nutrients are presented in three chapters – carnivorous plants, parasitic plants and specialised roots. An entire chapter is devoted to leaves, with an insight into how leaves may assist in protecting flowers and fruits from herbivores and seed-eaters. The book provides an ecological perspective on how the flora has evolved complex strategies to ensure species survival in the relatively harsh seasonal climate of a Mediterranean-type ecosystem.

- 268 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
1 Evolution and Diversity of the Flora
Land plants arose almost 500 million years ago (Ma). Those with a vascular system (lycopods) appeared at least 450 Ma, followed by the ferns with true leaves rather than scales at least 400 Ma. Seed-bearing plants originated about 430 Ma giving rise to the gymnosperms about 330 Ma, including the cone-bearing cycads in one clade (phylogenetic group) and the conifers in another.
The latest research based on rates of change in DNA structure (the molecular clock) puts flowering plants arising at least 134 Ma but it may have been up to 248 Ma depending on the true affinity of different fossils and the accepted rates of molecular change and whether a group is only recognised once it starts to separate into its daughter groups (called the crown) or at its base where it remains undivided or only a single-ancestor can still be identified at the molecular level (Clark et al., 2011). This puts angiosperm origins in the Permian Period, or as early as the Ordivician, much earlier than once thought. Nevertheless, once the angiosperms appeared they gradually outcompeted all previously existing groups of plants, certainly by the end of the Cretaceous, 65.5 Ma. This was apparently due to their rapid growth rates and superior colonizing ability. Speciation of the SouthWest angiosperm flora was particularly rapid during the Miocene to Pleistocene, a time of great climatic upheavals with a general drying and cooling trend from 15 Ma (Fig. 1.1), including a period of severe aridification 2−5 Ma (Dodson & Macphail, 2004). Many plant lineages that were once widespread within temperate Australia, now have separate taxa in the SouthWest and southeast/eastern Australia because of the biogeographical barrier imposed by aridification and the elevation of limestone sediments in the current Nullarbor Plain, beginning 14 Ma (Crisp & Cook, 2007).
1.1 A Radiating and Hyperdiverse Flora
Elements of the Australian flora were already present in the Upper Cretaceous, 90 Ma, mostly confined to rainforest dominated by conifers but also in fire-prone, sclerophyll pockets (see Fig. 1.5 – Proteaceae phylogeny). The large asteroid that struck the Earth in the Gulf of Mexico 65.5 Ma, causing an abrupt end to the Cretaceous Period, was an important event for the world’s biota, including the SouthWest. Within 4 million years (61−62 Ma), key taxa in the SouthWest flora such as Banksia (Proteaceae), Eucalytpus (including Corymbia) (Myrtaceae) and the Australian Restionaceae emerged (Fig. 1.1). All these groups are adapted to nutrient-impoverished soils and highly fire-tolerant. Eucalypts are at present the dominant overstorey species in temperate Australian woodlands and forests and they further enhanced the flammability of the flora through the aromatic oils in their leaves and stems though it is thought that initially they evolved to deter herbivores (Crisp et al., 2011). Eucalypts also have a distinctive epicormic resprouting structure (Chapter 2; Burrows, 2013) that remained unchanged throughout the subsequent Cenozoic Era to the present time (Crisp et al., 2011).
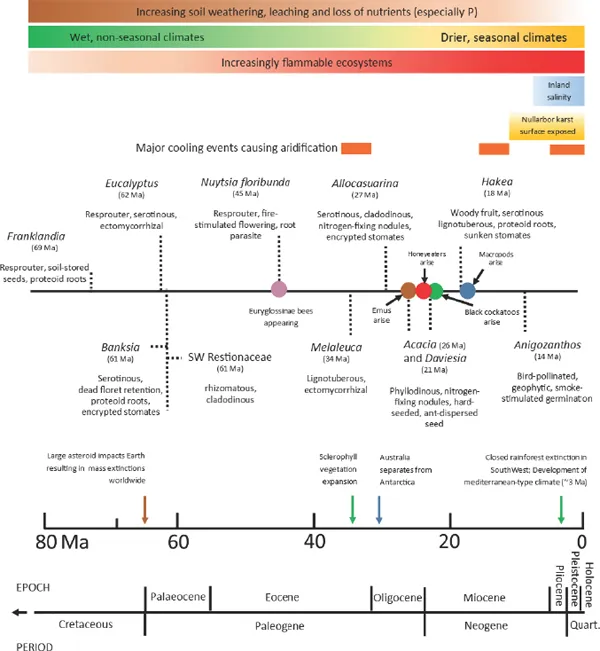
Fig. 1.1: Appearance of extant species-rich and iconic SouthWest taxa in Australia since the late Cretaceous Period compared with the occurrence of biotic and environmental agents of selection. Euryglossin bees and honeyeaters are pollinators of the flora (Chapter 8) whereas black cockatoos have influenced the evolution of woody-fruited species (e.g. Hakea) (Chapter 11). Geological time compiled from The Geological Society of America Geological Time Scale (2012, v. 4), and key events obtained from Byrne et al. (2014). Formation of Nullarbor Plain and aridification events since 35 Ma obtained from Crisp & Cook (2007). Information obtained from time-calibrated molecular phylogenies described in Crisp et al. (2004) [Allocasuarina]; Vidal-Russell & Nickrent (2008a) [Nuytsia]; Hopper et al. (2009) [Anigozanthos]; Prideaux & Warburton (2010) [Macropods]; He et al. (2011) [Banksia]; Crisp et al. (2011) [Eucalyptus, Melaleuca]; White et al. (2011) [Calyptorhynchinae cockatoos]; Miller et al. (2013) [Acacia]; Almeida et al. (2012) [Euryglossinae bees]; Litsios et al. (2013) [Restionaceae]; Baker et al. (2014) [emus]; Joseph et al. (2014) [honeyeaters]; Franklandia and Hakea from Fig. 1.5.
Tree root-parasites are basal within the hemiparasitic Santalales order and the best known in the SouthWest is the monotypic Nuystia floribunda (Loranthaceae) that arose 45 Ma during the Eocene (Fig. 1.1). This relict is unique among the Santalales in having wind-dispersed fruits (all others have succulent fruits that are a feature of rainforest species), precocious fire-stimulated flowering from fireblackened stems, tough, semi-terete leaves, and rhizomes reaching 100 m in length. These features suggest that at least parts of the SouthWest during the Eocene had a highly seasonal climate and were subject to frequent fire (Lamont & He, 2012). Parasitism in the Loranthaceae evolved from terrestrial to aerial 28 Ma, with rapid diversification occurring during the Oligocene, an epoch of global cooling and temperate woodlands and grasslands (Vidal-Russell & Nickrent, 2008a). Today Loranthaceae is dominated by aerial hemiparasites, though only Ameyma and Lysiana are present (and widespread) in the SouthWest (Chapter 5). Both genera are pollinated by honeyeaters (Meliphagidae) that originated 23.5 Ma and radiated strongly between 15 to 5 Ma (Joseph et al., 2014; Fig. 1.1). The frugivorous mistletoe bird (Dicaeum hirundinaceum, Dicaeidae) is a major seed disperser (Yan, 1993a), although this species did not colonise Australia until the Pleistocene, possibly as late as the Holocene (Reid, 1987), arriving from New Guinea and the surrounding islands (Nyári et al., 2009) and was already adapted to its mistletoe berry diet. This bird never reached Tasmania.
The Ecocene also saw the origin of solitary stingless bees in the Australian endemic subfamily Euryglossinae (Colletidae) (Fig. 1.1) and the Australian biodiverse subfamily Hylaeinae (48-54 Ma) (Almeida et al., 2012) that are important pollinators of small-flowered species in Australia. Emus (large flightless bird) (Dromaius novae-hollandiae: Dromaiidae) appeared 27 Ma (Fig. 1.1) and became important long-distance seed dispersers.
Major taxonomic groups were well established in the Australian flora by the time Antarctica split from Australia as it pulled north, beginning in the West in the early Eocene and finishing with Tasmania during the early Oligocene, leaving the flora to radiate in isolation from other continents, as the climate became colder and drier with Australia’s drift into the mid-latitudes. The Casuarinaceae (Allocasuarina, Casuarina and Gymnostoma) radiated about 47 Ma (Crisp, Cook & Steane, 2004), with the Allocasuarina/Casuarina clade developing about 39 Ma, with their leaves reduced to minute scales, encrypted stomates hidden within cladodinous grooves, and serotinous woody cones (Crisp & Cook, 2013). Speciation in Allocasuarina appears to have accelerated about 25 Ma dominating the expanding temperate sclerophyllous floras of the SouthWest and southeastern Australia (Crisp et al., 2004).
The greatest rates of speciation occurred during the long Miocene Epoch (Fig. 1.1), as the climate became drier and more seasonal. Rates of trait proliferation were highest in both directions at that time. For example, the rates of evolution of species with serotiny and non-serotiny, retention and shedding of dead florets and leaves, and clonality and non-clonality among the genus Banksia were greatest then (He et al., 2011). This indicates that a great range of habitats and climate extremes was present at that time and that the Miocene was an important period in the evolution of the SouthWest flora (and Australia generally). The only exception among banksias was the thickening of the mantle of dead florets that finally covered the fruits that peaked in the Pliocene (2−5 Ma), either as a response to increasing reliance on fire to melt the resin sealing the fruits or to improve crypsis from the increasing presence of granivorous black cockatoos (C...
Table of contents
- Cover
- Title
- Copyright
- Prologue
- Acknowledgments
- Contents
- 1 Evolution and Diversity of the Flora
- 2 Fire Adaptations
- 3 Drought Responses
- 4 Carnivorous Plants
- 5 Parasitic Plants
- 6 Specialised Nutrient Uptake Mechanisms
- 7 Pollination Strategies
- 8 Pollination Syndromes
- 9 Leaf Properties
- 10 Seed Release and Dispersal Mechanisms
- 11 Seed Storage, Germination and Establishment
- Bibliography
- About the Authors
- List of Figures
- List of Tables
- Index
- Backcover
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Plant Life of Southwestern Australia by Philip Groom, Byron Lamont in PDF and/or ePUB format, as well as other popular books in Ciencias biológicas & Ecología. We have over 1.5 million books available in our catalogue for you to explore.