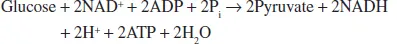![]()
Chapter 1
Glycolysis and Its Metabolic Engineering Applications
Jian Wang* and Yajun Yan†,‡
*College of Engineering, University of Georgia
Athens, GA 30602, USA
†BioChemical Engineering Program, College of Engineering
University of Georgia, Athens, GA 30602, USA
1.1.Introduction
Microorganisms play a pivotal role in the degradation of complex carbon sources in nature and could survive in different conditions due to the genetically coined cellular metabolism. Cellular metabolism of microorganisms is defined as the sum of biochemical processes that involves the conversion of environmental nutrients into simple biosynthetic building blocks and energy in the cells. Within these metabolic processes, catabolism is responsible for breakdown of complex molecules into simple molecules, producing energy, reducing power, and precursor intermediates for other metabolic processes like anabolism. The central catabolic pathways include Embden–Meyerhof–Parnas (EMP) pathway, pentose phosphate (PP) pathway, Entner–Doudoroff (ED) pathway, and the tricarboxylic acid (TCA) cycle. The EMP pathway, also known as glycolysis pathway, is one of the most ancient metabolic pathways and occurs, if at least in part, in nearly all organisms.1 Glycolysis pathway is an oxygen-independent metabolic pathway that comprises the initial steps required for the metabolism of carbohydrates like glucose. Its function is essential for cell growth because it generates precursor metabolites from central catabolic pathways that are the starting building blocks for macromolecules and other small molecules. Thus, glycolysis pathway is a gatekeeper pathway for carbohydrate utilization and cell survival. This chapter mainly focuses on the fundamental mechanisms and metabolic engineering applications of glycolysis pathway for the production of a plethora of value-added chemicals in the well-developed Escherichia coli.
1.2.The Fundamentals of Glycolysis in Biological Systems
1.2.1.Glycolysis Pathway
Glycolysis pathway is one of the main components of central metabolism that is regarded as an entry pathway for glucose utilization. The principal functions of glycolysis pathway are oxidation of hexoses like glucose to generate adenosine triphosphate (ATP), reductants like reduced nicotinamide adenine dinucleotide (NADH) and trioses like pyruvate. During this process, six of 12 precursor metabolites are supplied for biosynthesis (Table 1.1). The reactions of glycolysis pathway can be summarized as a reaction as follows:
Glycolysis pathway is a 10-step pathway and can be divided into two stages: the preparatory phase glucose to glyceraldehyde-3-phosphate (G-3-P) and the payoff phase (G-3-P to pyruvate) (Figure 1.1). The preparatory phase consists of the first five steps that include glucose phosphorylation, isomerization, group transfer, and cleavage reactions. Two moles of ATP are utilized and two moles of G-3-P are produced per mole of glucose. The payoff phase consists of subsequent five redox reactions that oxidize G-3-P to pyruvate, during which four moles of ATP and two moles of NADH are produced per mole of glucose. Glycolysis pathway-mediated glucose oxidation is incomplete and the final products (two pyruvates) still contain the bulk of the total energy initially in glucose. Complete oxidation of one mole of glucose will release 2,840 kJ of energy, whereas 94.8% of this energy is stored in two moles of pyruvate, which will be largely released during TCA cycle.2
Table 1.1. Twelve major precursor metabolites generated in central pathways.
Precursor metabolite | Metabolic pathway |
Glucose-6-phosphate (G-6-P) | Glycolysis |
Fructose-6-phosphate (F-6-P) | Glycolysis |
Triose phosphate | Glycolysis |
3-Phosphoglycerate (3-PG) | Glycolysis |
Phosphoenolpyruvate (PEP) | Glycolysis |
Pyruvate | Glycolysis |
Ribose-5-phosphate (R-5-P) | PP pathway |
Erythrose-4-phosphate (E-4-P) | PP pathway |
Acetyl-CoA | TCA cycle |
α-Ketoglutarate (α-KG) | TCA cycle |
Oxaloacetate | TCA cycle |
Succinyl-CoA | TCA cycle |
In E. coli, the assimilation and phosphorylation of glucose can be realized by the phosphotransferase system (PTS) or by glucokinase (encoded by glk). The PTS system, coupled with consumption of one mole of phosphoenolpyruvate (PEP) to pyruvate for each mole of internalized glucose, plays a major role in glucose transport and phosphorylation.3 The PTS system consists of sugar-non-specific protein components (Enzyme I, encoded by ptsI; Hpr, encoded by ptsH) and glucose-specific PTS permeases (Enzyme IIAglc, encoded by crr; Enzyme IIBCglc, encoded by ptsG). The phosphate group from PEP is transferred sequentially through Enzyme I, Hpr, Enzyme IIAglc, Enzyme IICglc, and IIBglc to glucose.3 Glucokinase can also produce phosphorylate glucose, but it is not indispensable for cell growth on glucose as the carbon source.4 Phosphoglucose isomerase (encoded by pgi) catalyzes the isomerization of glucose-6-phosphate (G-6-P) to fructose-6-phosphate (F-6-P), an essential step of the glycolysis pathway. Two 6-phosphofructokinase isozymes, Pfk I and Pfk II (encoded by pfkA and pfkB), do not share sequence similarity and catalyze the phosphorylation of F-6-P on the C1 carbon with the involvement of ATP consumption. More than 90% of the phosphofructokinase activity are attributed to Pfk I and only less than 5% are attributed to Pfk II.5 E. coli also contains two classes of fructose-1,6-bisphosphate aldolases, FbaA and FbaB (encoded by fbaA and fbaB), which catalyze a reversible aldol cleavage/condensation reaction of fructose 1,6-bisphosphate (FBP) during glycolysis and gluconeogenesis.6 The class II aldolase FbaA (metallo) utilizes a divalent metal ion zinc while the class I aldolase FbaB (Schiff base) utilizes a catalytic lysine residue (Lys236) to stabilize the catalytic intermediate. FbaA is required for glycolysis while FbaB most likely participates in gluconeogenesis as it is not expressed when grown on glucose but induced when grown on C3 carbon sources.7,8 In the glycolytic direction, FBP is cleaved to produce dihydroxyacetone phosphate (DHAP) and G-3-P. Triosephosphate isomerase (TpiA) (encoded by tpi) catalyzes isomerization between G-3-P and DHAP. G-3-P dehydrogenase A (encoded by gapA) catalyzes the reversible oxidative phosphorylation of G-3-P to 1,3-bisphosphoglycerate (1,3-BPG) in the presence of NAD+. In the glycolytic direction, two moles of NADH will be produced per mole of glucose consumed. Phosphoglycerate kinase (Pgk; encoded by gene pgk) is responsible for the reversible phosphorylation between 3-phosphoglycerate (3-PG) and 1,3-BPG. In the glycolytic direction, Pgk catalyzes the transfer of a phosphoryl group from 1,3-BPG to ADP, forming ATP and 3-PG. E. coli contains both a 2,3-bisphosphoglyerate-dependent phosphoglycerate mutase GpmA (encoded by gpmA) and a cofactor-independent phosphoglycerate mutase GpmM (encoded by gpmM), which catalyze the interconversion between 3-PG and 2-phosphoglycerate (2-PG). The GpmA enzyme has significantly higher specific activity than GpmM.9,10 Enolase (encoded by eno) catalyzes the interconversion of 2-PG and PEP. Two pyruvate kinases, PykA and PykF (encoded by pykA and pykF), are key and last enzymes of the glycolysis pathway, catalyzing the irreversible transfer of the phosphoryl group o...