Understanding animal andrology is fundamental to optimising genetic breeding traits in domestic and wild animals. This book provides extensive coverage of male reproductive biology, discussing the essentials of sperm production, harvest and preservation before covering the applications to a range of animals including cattle, horses, pigs, small ruminants, camelids, cats and dogs, poultry and exotic species. It also examines the laboratory procedures that provide the basis of general fertility research.

eBook - ePub
Animal Andrology
Theories and Applications
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Animal Andrology
Theories and Applications
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Part I
Animal Andrology Theories
1 Semen and its Constituents
Brian P. Setchell*
University of Adelaide, Adelaide, South Australia
University of Adelaide, Adelaide, South Australia
Introduction
Semen, the material that is emitted from the penis at ejaculation, comprises a cellular component, the spermatozoa, and a liquid phase, the seminal plasma. The volume of semen in a single ejaculate varies widely among the domesticated mammals, from about 1 µl in sheep and goats to as much as half a litre in pigs. The density of spermatozoa also varies, being much higher in those species with small ejaculates and lower in those ejaculating large volumes.
Semen is composed of secretions of the ampulla of the ductus deferens, and of the accessory glands, seminal vesicles and prostate, as well as fluid and spermatozoa from the cauda epididymis. Individual components may have different origins.
Composition of Semen
Spermatozoa
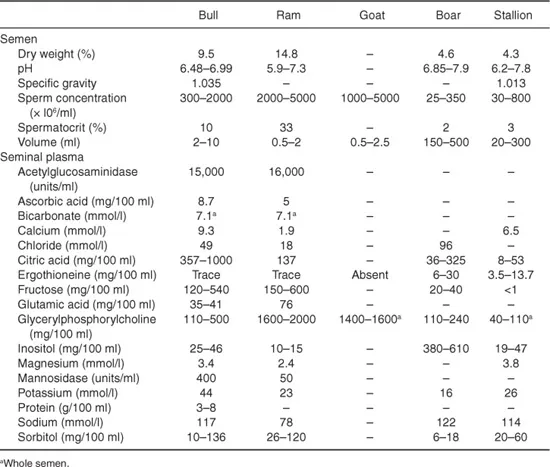
The fraction of the semen made up by spermatozoa is known as the spermatocrit, and ranges from more than 30% in sheep to less than 2% in pigs (Table 1.1).
A large amount of information is now available about the structure of the spermatozoa, and for details the reader is referred to the many detailed reviews on this topic (Bishop and Walton, 1960; Phillips, 1975; Bedford and Hoskins, 1990; Gage, 1998; Bedford, 2004; Eddy, 2006). In brief, the spermatozoa of the domestic mammals have spatulate heads containing the nuclear DNA, with an acrosome covering the anterior pole, attached by a specialized neck structure to a midpiece and tail. The midpiece consists of a helix of mitochondria surrounding the central two and surrounding nine fibres, which extend into the tail. The sperm of the domestic mammals are relatively small, at least when compared with those of most rodents, and are similar in size and structure to human sperm. The sperm of most murid rodents are much larger and quite different in shape, being falciform or hook shaped, with the acrosome over one side of the head.
Other cells
As well as spermatozoa, white blood cells (WBC) are often found in semen. In humans, more than 10 WBC/ml semen is often associated with infertility (Wolff, 1995), although this view is now not universally accepted (Aitken and Baker, 1995; Lackner et al., 2010; Tremellen and Tunc, 2010; Henkel 2011). In domestic mammals, WBC are often present in small numbers in semen, although there appears to be no relationship between their numbers and abnormalities of the sperm (Sprecher et al., 1999; Sutovsky et al., 2007; Alghamdi et al., 2010).
Table 1.1. Some details of the composition of the semen of the domestic animals. Based on data from Mann, 1964; Mann and Lutwak-Mann, 1981. Reproduced from Setchell, 1991, with permission from Elsevier.
Carbohydrates
One of the most remarkable features of semen is that the predominant reducing sugar is not glucose, as in blood, but fructose (Mann 1946a,b), a sugar more usually found in plants. Small amounts of glucose are also present, and boar semen in particular contain large concentrations of inositol, but less fructose than semen from bulls or rams (Mann, 1951). Stallion semen also contains inositol and lower concentrations of fructose (Baronos, 1951; Mann et al., 1963), and other compounds of inositol are also present in some species (Seamark et al., 1968). Fructose in bulls and rams originates in the seminal vesicles, with some from the ampulla, but in the stallion, most comes from the ampulla. Inositol is secreted in the seminal vesicles (Mann and Lutwak-Mann, 1981).
Both glucose and fructose can be utilized by sperm, either by oxidation or glycolysis, although the Michaelis constant (Km) for glucose is much lower than that for fructose (see Ford and Rees, 1990). The mitochondria, in which oxidative phosphorylation occurs, are arranged as a helix around the midpiece of the sperm, whereas the glycolytic enzymes are concentrated in the principal piece of the tail, while some are bound to the fibrous sheath of the flagellum. However, it is unlikely that glycolysis alone could generate enough ATP for full motility, and while diffusion from the mitochondria may be sufficient in smaller sperm, in larger sperm it is likely that an adenylate kinase shuttle is involved in moving ATP from the mitochondria to the flagellum (Ford, 2006; Miki, 2007; Storey, 2008; Cummins, 2009). There is evidence for the occurrence in sperm of specific glucose transporters that can transport both glucose and fructose (Purcell and Moley, 2009).
Proteins, amino acids and other nitrogen-containing compounds
Seminal plasma contains a variety of proteins and peptides, the total concentration being somewhat less than that in blood plasma (Mann and Lutwak-Mann, 1981). Seminal plasma proteins are derived from the epididymis and the accessory glands, and are involved in several essential steps preceding fertilization, including capacitation, establishment of the oviductal sperm reservoir, modulation of the uterine immune response, sperm transport in the female tract and gamete interaction and fusion (Calvete et al., 1994; Topfer-Petersen et al., 2005; Karekoski et al., 2011).
Some proteins are higher in the semen of fertile bulls, whereas others are more abundant in the semen of bulls of lower fertility (Killian et al., 1993; Bellin et al., 1998; Brandon et al., 1999). In stallions, the abundance of some proteins (kallikrein-1E2, clusterin and seminal plasma proteins 1 and 2 (SP1 and 2) are negatively related to fertility, whereas cysteine-rich secretory protein 3 (CRISP3) is positively related (Novak et al., 2010).
Other proteins are involved in sperm–egg interactions and cell cycle regulation (Gaviraghi et al., 2010). Identified proteins include leptin and insulin-like growth factor I (IGF-I; Lackey et al., 2002) and phospholipidbinding proteins involved in sperm membrane lipid modification during capacitation (Manjunath and Therien, 2002). Seminal plasma from stallions contains SSP-7 (stallion seminal protein 7, also known as horse seminal protein 7 – HSP-7), a member of the spermadhesin protein family that is involved in the sperm binding to the zona pellucida of the oocyte (Reinert et al., 1997), and there are also heparin-binding proteins, which modulate capacitation (Miller et al., 1990; Nass et al., 1990; Bellin et al., 1994). Other proteins inhibit in vitro and cooling-induced capacitation (Vadnais and Roberts, 2010) and the ability of sperm to penetrate zona-free oocytes (Henault et al., 1995; Henault and Killian, 1996), as well as sperm transport and elimination (Troedsson et al., 2005), sperm longevity (Karekoski and Katila, 2008) and storage in the oviduct (Gwathmey et al., 2006). Seminal plasma from pigs contains high concentrations of transforming growth factor β (TGF-beta), an important immune deviating agent (Robertson et al., 2002).
Seminal plasma also contains considerable concentrations of free amino acids, particularly glutamic acid in rams and bulls (Setchell et al., 1967; Brown-Woodman and White, 1974) and hypotaurine in boars (Van der Horst and Grooten, 1966; Johnson et al., 1972). Hypotaurine may be important in preventing damage to sperm by reactive oxygen species (Alvarez and Storey, 1983; Bucak et al., 2009).
There are also appreciable concentrations of carnitine in the seminal plasma of rams (Brooks, 1979), bulls (Carter et al., 1980) and stallions (Stradaioli et al., 2004). This substance is involved in fatty acid transport in other tissues, but that present in semen is largely derived from the epididymis (Hinton et al., 1979). Boar semen also contains ergothioneine, the betaine of thiolhistidine, a sulfur-containing reducing base, which comes mainly from the seminal vesicle (Mann and Leone, 1953); it is also present in stallion semen, but in this species, it originates largely from the ampulla (Mann and Lutwak-Mann, 1963).
Semen and seminal plasma from rams, bulls, goats, boars and stallions were found to contain considerable amounts of glycerophosphorylcholine, which originates largely from the epididymis (Dawson et al., 1957; Brooks 1970), as well as glycerylphosphorylinositol.
Lipids
Semen contains considerable amounts of lipid, both neutral lipids and phopholipids, most of which is in the spermatozoa (Hartree and Mann, 1959). In ram semen, the most abundant phospholipid is choline plasmalogen (also known as phosphatidalcholine), whereas in boars, it is lecithin (also known as phosphatidylcholine) and in bull sperm, the two phospholipids are present in approximately equal amounts (see Mann and Lutwak-Mann, 1981). One remarkable feature of these phospholipids is their high concentration of highly unsaturated fatty acids, 22 carbons in length, with six double bonds (22:6) in rams and bulls and five double bonds (22:5) in boars (Johnson et al., 1969; Poulos et al., 1973; Evans and Setchell, 1978). These constituent fatty acids are particularly susceptible to damage from reactive oxygen species. The phospholipids may also be important precursors of platelet activating factor (PAF), which is probably involved in sperm motility, the acrosome reaction and fertilization, and which is found in bull and boar sperm (Parks et al., 1990; Roudebush and Diehl, 2001). Seminal plasma from bulls and stallions contains an acetylhydrolase, which may play a role in regulating autocrine or paracrine functions of PAF (Parks and Hough, 1993; Hough and Parks, 1994).
Semen also contains appreciable concentrations of steroids. In bull semen, the concentrations of several steroids, including progesterone, dihydrotestosterone, androstanediols and oestrogens are much higher than in blood plasma. The oestrogens appear to come from the prostate, whereas the other steroids originate from the epididymis. Testosterone is present in seminal plasma at about the same concentration as in blood plasma, much less than in the rete testis fluid leaving the testis (Ganjam and Amann, 1976).
Prostaglandins were discovered in the 1930s, and were so named because it was thought that they came from the prostate, but in fact they originate largely from the seminal vesicle in rams and bulls. They occur in smaller concentrations in the testis and epididymis (Voglmayr, 1973; Kelly, 1978).
Function of Semen
Transport of spermatozoa
An obvious function of the semen is the transport of the spermatozoa into the female reproductive tract at mating. The site of deposition varies according to species; it is deposited into the vagina in cattle and sheep, but directly into the uterus in pigs and to some extent in horses. (Anderson, 1991). Movements of the female tract – which are probably important in moving the spermatozoa from the site of deposition to that of fertilization – are probably influenced by some seminal constituents, in particular by prostaglandins (Kelly, 1978).
Metabolism of spermatozoa
It has been argued that the spermatozoa are in contact with the seminal plasma for too short a time for metabolism of their constituents to be of major importance, but several hours can elapse between mating and fertilization, so some utilization of metabolites, particularly of sugars, should be possible. However, the sperm probably also utilize cellular constituents, particularly lipids, during this time.
Effects of seminal plasma on the female reproductive tract
The possible involvement of proteins in the seminal plasma in the establishment of oviductal sperm reserves, the capacitation of sperm and the processes of fertilization, including binding to and penetration of the zona pellucida of the oocyte, has already been mentioned. It should be remembered, though, that the conceptus must also be protected from maternal immune attack...
Table of contents
- Cover Page
- Title Page
- Copyright Page
- Contents
- Contributors
- Preface
- Part I. Animal Andrology Theories
- Part II. Animal Andrology Applications
- Index
- Color Plates
- Footnotes
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Animal Andrology by Peter J Chenoweth, Steven P. Lorton, Peter J Chenoweth,Steven Lorton, Peter J Chenoweth, Steven Lorton in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.