Emerging and re-emerging pathogens pose several challenges to diagnosis, treatment, and public health surveillance, primarily because pathogen identification is a difficult and time-consuming process due to the "novel" nature of the agent. Proper identification requires a wide array of techniques, but the significance of these diagnostics is anticipated to increase with advances in newer molecular and nanobiotechnological interventions and health information technology.
Human Emerging and Re-emerging Infections covers the epidemiology, pathogenesis, diagnostics, clinical features, and public health risks posed by new viral and microbial infections. The book includes detailed coverage on the molecular mechanisms of pathogenesis, development of various diagnostic tools, diagnostic assays and their limitations, key research priorities, and new technologies in infection diagnostics. Volume 1 addresses viral and parasitic infections, while volume 2 delves into bacterial and mycotic infections.
Human Emerging and Re-emerging Infections is an invaluable resource for researchers in parasitologists, microbiology, Immunology, neurology and virology, as well as clinicians and students interested in understanding the current knowledge and future directions of infectious diseases.
Trusted by 375,005 students
Access to over 1 million titles for a fair monthly price.
Chapter 1 Pathogenesis of the Old World Arenaviruses in Humans
Igor S. Lukashevich
Department of Pharmacology and Toxicology, School of Medicine, Center for Predictive Medicine, NIH Regional Bio-containment Laboratory, University of Louisville, Louisville, USA
1.1 The Old World arenaviruses: taxonomic and zoonotic introduction
Arenaviruses represent a fast-growing group of rodent-borne viruses (see Notes in the Proofs) which are an example of how environmental changes disrupt the natural animal virus–host balance and result in unexpected diseases. In the wild, arenaviruses exist as chronic infections in specific rodent hosts. This provides ideal conditions for competition within viral quasispecies for improved adaptation to the host. Analysis of arenavirus phylogeny and rodent cytochrome-b sequences provides examples of coevolution of arenaviruses with their rodent hosts. Based on their antigenic properties and geographic distribution approximately two dozen arenaviruses discovered so far are placed into two groups: the Old World (OW) or Lassa–LCMV complex and the New World (NW) or Tacaribe complex (Salvato et al., 2012). In general, the OW viruses are hosted by rodents of the family Muridae, subfamily Murinae. The NW arenaviruses are associated with rodents of the subfamily Sigmodontinae which are divided into the North and South American lineages. Phylogenetically the NW arenaviruses are further divided into clades A, B, C, and a recombinant A/B clade.
Lymphocytic choriomeningitis virus (LCMV), the prototypic arenavirus that belongs to the OW group, is hosted by the house mouse (Mus musculus). The virus is distributed worldwide and can cause aseptic meningitis or meningoencephalitis in immunocompetent children and adults. Recent studies indicate that LCMV is also an under-recognized cause of congenital infection and neurological disease in the fetus and newborns (Bonthius, 2012; Laposova et al., 2013). In addition, LCMV and/or LCMV-like viruses have been also associated with clusters of fatal illness among tissue transplant recipients (Waggoner et al., 2013). All other human pathogens from the Arenaviridae cause viral hemorrhagic fevers (VHFs).
The VHFs encompass a diverse group of viral infectious diseases of animals and humans caused by distinct RNA viruses from four major families. In humans these viruses can induce severe life-threatening diseases, with fever, malaise, hemorrhage, and hypovolemic shock resembling bacterial LPS (lipopolysaccharide)-induced sepsis-like shock at terminal stage of the diseases. The VHFs may occur as isolated case(s), such as imported case(s) from endemic areas, or may cause devastating lethal outbreaks (Goeijenbier et al., 2013; Ippolito et al., 2012; Moraz and Kunz, 2011; Vela, 2010).
The Arenaviridae family contains the largest number of virus species causing VHFs. Lassa virus (LASV) is the most prominent human pathogen of the Arenaviridae responsible for several hundred thousand infections and death of thousands of patients with Lassa fever (LF) annually in West Africa (Fichet-Calvet and Rogers, 2009; Richmond and Baglole, 2003). Recently isolated in South Africa, LuJo virus (LUJV) will be a novel OW species (Briese et al., 2009). All pathogenic NW arenaviruses causing South American VHFs belong to clade B and include Junin virus (JUNV), Machupo virus (MACV), Guanarito virus (GTOV), Sabia virus (SABV), and the recently discovered Chapare virus (CHAPV) (Salvato et al., 2012).
In addition to arenaviruses, causative agents of VHFs were found among Bunyaviridae (e.g., Crimean Congo HF, CCHF; hemorrhagic fever with renal syndrome, HFRS; Rift Valley fever, RVF), Filoviridae (Ebola and Marburg HFs), and Flaviviridae (e.g., dengue HF; yellow fever, YF). With recent discovery of Bas-Congo virus isolated from human HF outbreak in Congo (Grard et al., 2012), Rhabdoviridae family, comprising the most diverse and widely distributed viruses in nature (Dietzgen et al., 2012), can be added to this list.
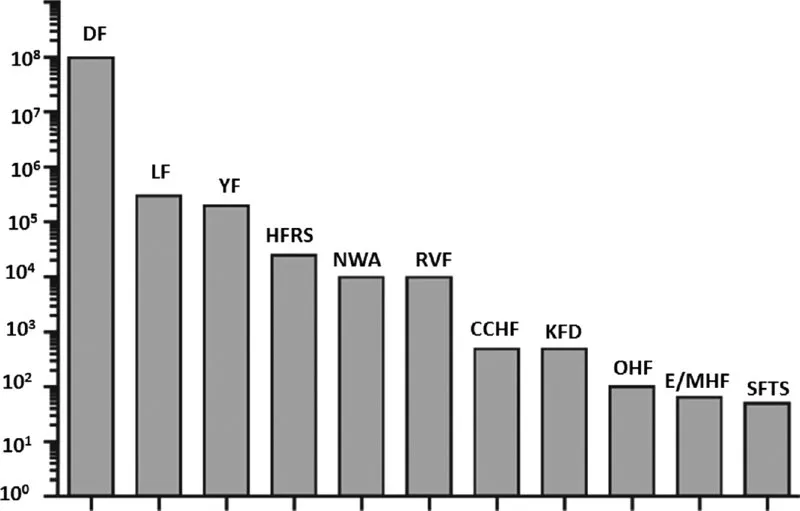
VHFs are predominantly zoonotic infections existing in certain geographic areas. According to the most recently published estimates (Falzarano and Feldmann, 2013), more than a third of the world's population live in areas that are at risk for VHFs. As shown in Figure 1.1, the most prevalent VHFs are dengue HF, LF, and YF. Climate changes and urbanization affect behavior of viral vectors (e.g., rodents, ticks, mosquitoes). These factors, together with substantial improvements in diagnosis and virus detection, suggest that novel viruses, including human pathogens, will continue to emerge in the coming years.
Fig. 1.1 Estimated global burden of viral hemorrhagic fevers (approximately 500,000 cases per year): DF, Dengue HF and Dengue shock syndrome; LF, Lassa Fever; YF, Yellow fever; HFRS, HF fever with renal syndrome; NWA, HF caused by OW and NW arenaviruses; RVF, Rift Valley fever; CCHF, Congo-Crimean HF; KFD, Kyasanur forest disease; OHF, Omsk HF; E/MHF, Ebola and Marburg HF; SFTS, severe fever with thrombocytopenia syndrome. From Falzarano and Feldmann, 2013. With permission from Elsevier.
Currently, only six OW arenavirus species have been recognized by the International Committee for Taxonomy of Viruses (ICTV) (Salvato et al., 2012). Among them, two viruses, LCMV and LASV, are human pathogens, and others, Mopeia (MOPV), Morogoro (MORV), Mobala (MOBV), and Ippy (IPPYV), are non-pathogenic virus species. To be defined as a novel arenavirus species the putative virus must meet the following criteria: (i) association with a host species; (ii) circulation in a defined geographic area; (iii) human disease association; (iv) lack of cross-neutralization activity with genetically related arenaviruses; and (v) significant (>12%) amino acid difference from other arenavirus species (Salvato et al., 2012). During the last decade, several arenaviruses have been isolated, mostly in Africa. Some of them have been identified as putative new species of the OW group of arenaviruses.
In 2008, LUJV caused a HF outbreak with unprecedentedly high case-fatality rate in Lusaka (Zambia) and Johannesburg (South Africa) (Briese et al., 2009; Paweska et al., 2009). Phylogenetic analysis showed that LUJV is a novel OW species and branches off the ancestral node of the OW arenaviruses. Screening of Mastomys natalensis trapped in Zambia, where the first patient infected with LUJV was identified, resulted in the isolation of Luna virus (LUNV) genetically related to non-pathogenic MOBV (Ishii et al., 2011).
LCMV-like Dandenong virus (DANV) was isolated from one of the three transplant recipients who received visceral organs (liver and kidneys) from the same donor from the former Yugoslavia. All three recipients died from febrile illness (Palacios et al., 2008). Phylogenetic analysis revealed that DANV is most closely related to LCMV. One more virus, named Kodoko virus (KODV), related to but distinct from LCMV, was isolated in Guinea from Mus (Nannomys) minutoides (Albariño et al., 2010; Lecompte et al., 2007).
In 1985, Merino Walk virus (MWV), a proposed novel tentative species, was isolated from a novel rodent host Myotomys unisulcatus collected at Merino Walk at Eastern Cape (South Africa) (Palacios et al., 2010). Full-length genomic sequence revealed that MWV is distantly related to sublineage MOBV-MOPV-IPPYV with the lowest amino acid sequence divergence, 31.4%, to MOPV.
In several field studies, screening of small mammals in Africa with advanced molecular virology tools resulted in detection of potentially novel OW species based only on RNA sequence analysis, without isolation of replication-competent virus. In Tanzania, search for rodent hosts for Morogoro virus (MORV) using RT-PCR targeting the arenavirus large (L) gene (see sub-section 1.2.1), revealed a novel arenavirus sequence from blood samples of Lemniscomys rosalia (de Bellocq et al., 2010). A portion of the L gene sequence (320 bp) encoded the amino acid sequence which was clustered with IPPYV but had 17.3% divergence (e.g., amino acid divergence between LASV and MOBV is 14.5%), indicating that the L sequence found in L. rosalia appears to be associated with a new species of OW arenavirus.
Extensive screening for novel arenaviruses in Cote d'Ivoire using pan-OW arenavirus RT-PCR primers targeting small (S) and L RNA genome segments identified unique sequences of putative novel Menekre virus (MENV) and Gbagroube virus (GBAV), the sequences of which were detected in Hylomyscus sp. and Mus (Nannomys) setulosus, respectively (Coulibaly-N'Golo et al., 2011). Notably, the GBAV sequence was closely related to LASV, while the MENV sequence clustered with IPPYV-MOBV-MOPV. Detection of LASV-like sequences in Mus setulosus suggests that co-evolution of African arenaviruses and their hosts can potentially include host-switching events, predicting isolation of novel arenavirus species in the future.
Recently, a group of highly diverse novel viruses distantly related to arenaviruses but also to filoviruses was isolated from snakes with fatal boid inclusion body disease (BIBD) (Bodewes et al., 2013; Hetzel et al., 2013; Koellhoffer et al., 2014; Stenglein et al., 2012). These viruses can be potentially placed in a novel taxonomic entity within the Arenaviridae with putative name BIBD-associated arenaviruses. Preliminary phylogenetic analysis based on conservative RNA polymerase motifs showed that snake-borne arenaviruses shared ancestry with NW and OW arenaviruses. While taxonomic status of these viruses is not defined, it is obvious that phylogenic tree of the Arenaviridae will be significantly changed in the nearest future (see Notes in the Proofs). Importantly, isolation of BIBD arenaviruses from non-mammal hosts indicates that these viruses can infect very broad range of species with unpredictable pathogenic potential for humans.
This chapter focuses on the OW arenavirus human pathogens, LCMV, LASV, and LUJV. LASV has the highest human impact of any VHF (with the exception of Dengue HF), and the aim of this chapter is to summarize our current knowledge on pathogenesis of LASV infection in experimentally infected non-human primates and on natural LF in humans. Although pathogenic and non-pathogenic arenaviruses shared many similarities in terms of molecular structure and mechanism of replication, recent studies revealed clear differences at the early stage of the host responses even between closely related arenaviruses with different pathogenic potential (Djavani et al., 2007; Malhotra et al., 2013; Zapata et al., 2013b). It indicates that key events determining the infection outcome significantly precede viremia, before release of newly synthesized virions in circulating blood. Detection of “molecular signatures” discriminating pathogenic versus non-pathogenic arenavirus infections will improve diagnosis and treatment of patients.
1.2 Genome structure
1.2.1 Arenavirus genome structure
Purified and negatively stained LASV virions have typical arenavirus morphology and appear as round or oval particles with a mean diameter of approximately 110–130 nm (Figure 1.2). Internal granules within virions are morphologically similar to host ribosomes, and RNA isolated from purified LASV contains variable amount of host cell 28S and 18S ribosomal RNAs i...
Table of contents
Cover
Title page
Copyright
Dedication
List of Contributors
Preface
Acknowledgments
About the Editor
Chapter 1 Pathogenesis of the Old World Arenaviruses in Humans
Chapter 2 Pathogenesis of New World Arenaviruses in Humans
Chapter 3 Pathogenesis of Emerging and Novel Bunyaviruses in Humans
Chapter 4 Pathogenesis of Rift Valley Fever in Humans
Chapter 5 Pathogenesis of Hantavirus Infections
Chapter 6 Molecular Pathogenesis of Japanese Encephalitis Virus Infection
Chapter 7 Dengue Virus Infection in Humans: Epidemiology, Biology, Pathogenesis, and Clinical Aspects
Chapter 8 Pathogenesis of West Nile Virus in Humans
Chapter 9 Overview on Chikungunya Virus Pathogenesis
Chapter 10 Nipah Virus Infections in Humans
Chapter 11 Pathogenesis of Hendra Virus in Humans
Chapter 12 Pathogenesis of Rotavirus in Humans
Chapter 13 Pathogenesis of Papillomaviruses in Humans
Chapter 16 Pathogenesis of Toxoplasma gondii in Humans
Chapter 17 Pathogenesis of Human African Trypanosomiasis
Chapter 18 Pathogenesis of Leishmaniasis in Humans
Chapter 19 Pathogenesis of Chagas Disease in Humans
Chapter 20 Pathogenesis of Cryptosporidium in Humans
Chapter 21 Pathogenesis of Malarial Parasites in Humans
Chapter 22 Pathogenesis of Trichomonas vaginalis in Humans
Chapter 23 Loa loa Pathogenesis in Humans
Chapter 24 Nematode Larva Migrans
Chapter 25 Pathogenesis of Human Schistosomiasis
Chapter 26 Vector-Borne Parasitic Zoonotic Infections in Humans
Index
List of Contributors
Preface
Acknowledgments
About the Editor
Chapter 27 Pathogenesis of Haemophilus influenzae in Humans
Chapter 28 Campylobacter jejuni: Molecular Mechanisms and Animal Models of Colonization and Disease
Chapter 29 Pathogenesis of Francisella tularensis in Humans
Chapter 30 Pathogenesis of Yersinia pestis in Humans
Chapter 31 Pathogenesis of Legionella pneumophila in Humans
Chapter 32 Pathogenesis of Kingella kingae Infections
Chapter 33 Pathogenesis of Helicobacter pylori in Humans
Chapter 34 Pathogenesis of Chlamydia trachomatis in Humans
Chapter 35 Pertussis or Whooping Cough
Chapter 36 Pathogenesis of Human Ehrlichioses
Chapter 37 Neisseria gonorrhoeae: The Pathogen, Diagnosis, and Antimicrobial Resistance
Chapter 38 Pathogenesis of Corynebacterium diphtheriae and Corynebacterium ulcerans
Chapter 39 Pathogenesis of Staphylococcus aureus in Humans
Chapter 40 Pathogenesis of Listeria monocytogenes in Humans
Chapter 41 Anthrax Infection
Chapter 42 Pathogenesis of Streptococcus in Humans
Chapter 43 Pathogenesis of Clostridium botulinum in Humans
Chapter 44 Mycobacterium ulcerans and Buruli Ulcer
Chapter 45 Challenges Associated with Diagnostics, Drug Resistance, and Pathogenesis of Mycobacterium tuberculosis
Chapter 46 Pathogenesis of Orientia tsutsugamushi Infection in Humans
Chapter 47 Lyme Borreliosis
Chapter 48 NDM-Type Carbapenemases in Gram-Negative Rods
Chapter 49 Pathogenesis of Cryptococcosis in Humans
Chapter 50 Candida albicans: Clinical Relevance, Pathogenesis, and Host Immunity
Chapter 51 Pathogenesis of Pneumocystis jirovecii Pneumonia
Chapter 52 Pathogenesis of Aspergillosis in Humans
Index
EULA
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Human Emerging and Re-emerging Infections by Sunit Kumar Singh in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Microbiology. We have over one million books available in our catalogue for you to explore.